ООО "ПРОПИОНИКС"
| пн-пт с 09:00 до 18:00 | |
Cheeses With Propionic Acid Fermentation
СЫРЫ С ПРОПИОНОВОКИСЛЫМ БРОЖЕНИЕМ

Рисунок 35.1 Выбор сыров с пропионовокислым брожением

Cheese (Fourth Edition), Academic Press, 2017, Pages 889-910
|
СОДЕРЖАНИЕ
|
|
|
|
|
Пропионовокислое брожениие в сыре приводит к появлению характерных глазков и орехового и слегка сладковатого вкуса; обычно это достигается контролируемым образом с помощью культуры отобранных пропионовокислых бактерий (ПКБ). Однако естественная пропионибактериальная флора из сырого молока отличается большим штаммовым разнообразием и может вызывать спонтанное брожение в сырах, произведенных на основе такой флоры. В результате количество и размер глазков могут значительно варьироваться, а также могут часто встречаться трещины и расслоения. Применение культуры отобранных ПКБ позволяет добиться более регулярного формирования глазков и вкуса в результате контролируемого пропионовокислого брожения. Такие сорта сыра часто объединяют под общим термином "сыры швейцарского типа" (McSweeney et al., 2004). Тело и текстура таких сыров соответствуют твердым или полутвердым сырам. Изначально они производились как твердые сыры в Эмментале, долине в Швейцарии. Эмменталь PDO - это, пожалуй, самый известный сыр швейцарского типа, который берет свое начало в двенадцатом веке. В настоящее время во многих странах производится большое количество сыров швейцарского типа (рис. 35.1). Их можно разделить на следующие группы:
A. Сыры Эмменталь с защищенным наименованием происхождения (PDO) или защищенным географическим указанием (PGI)
"Emmentaler" PDO (Швейцария), "Allgäuer Emmentaler" PDO (Германия), "Emmental de Savoie" PGI (Франция) и "Emmental français est-central" PGI - вот сорта, представляющие эту группу сыров эмментальского типа. Продукт PDO должен быть полностью произведен в конкретном регионе традиционным способом, благодаря чему он приобретает уникальные свойства (European Commission, 2014). Этот процесс включает в себя производство сырого молока, превращение его в сыр и созревание сыров. Обычно несколько небольших молочных заводов в конкретном регионе производят эмментальские сорта с маркировкой PDO или PGI в соответствии с государственно-правовыми спецификациями; для обеспечения высшего качества таких сыров проводится оценка качества независимыми экспертами. Например, стандарт для Emmentaler PDO требует производства сыра без добавок и с минимальной обработкой в течение 24 часов после доения из свежего местного сырого молока от коров, которых кормили рационом с высоким содержанием грубых кормов (>70%) и без силоса для достижения высоких показателей качества молока, содержащего менее 25 спор ферментирующих лактат клостридий на литр. Сырные круги должны быть диаметром от 0,80 до 1,00 м, весить от 75 до 120 кг, иметь содержание жира в сухом веществе от 45,0% до 54,9% (полный жир) и максимальное содержание воды 380 г/кг (твердый сыр). Сухая натуральная корка должна иметь цвет охры или желтого цвета, а тело сыра должно быть эластичным, неклейким, слегка зернистым, иметь цвет от слоновой кости до бледно-желтого цвета и содержать отверстия преимущественно диаметром 2–4 см. Отверстия или "глазки" должны быть круглыми, четко очерченными и равномерно распределенными по всей поверхности. Выдержка сыра происходит в течение периода, варьирующегося от 5 месяцев до года и более. Сыр имеет мягкий, ореховый, сладкий вкус, который может усиливаться в процессе выдержки, и не соленый (Anonymous, 2012; Bisig et al., 2010). В отличие от этого, требования к продуктам PGI менее строгие, особенно в отношении происхождения сырья.
B. Эмментальский сыр в соответствии со Стандартом Кодекса
Типовые эмментальские сыры производятся в больших количествах по всему миру как твердые сыры в соответствии с гораздо менее строгим стандартом Codex alimentarius: 269-1967 для эмментальского сыра (Codex, 2014). В основном они производятся из пастеризованного молока и созревают недолго, но не менее 6 недель. Глазки, от скудных до обильных, должны быть размером от вишни до грецкого ореха и варьироваться от 1 до 5 см в диаметре. Допускается наличие нескольких пустот и трещин. Содержание воды может достигать 400 г/кг, что на 20 г/кг больше, чем для сыров PDO и PGI. Допускаются добавки, такие как красители, регуляторы кислотности и консерванты; 24 из них содержатся в списке Кодекса (Codex, 2014). Эмментальские сыры этой категории сегодня производятся во многих странах, в том числе во Франции, Германии, Финляндии и США.
C. Полутвердые сыры, изготовленные с использованием мезофильной закваски и ПКБ (Goutaler)
По открытости и вкусу Ярлсберг (Норвегия), Маасдам (Нидерланды), Самсое (Дания), Альпсберг и Фельсберг (Германия), Греве (Швеция) несколько похожи на Эмменталер PDO и Эмменталь (рис. 35.1); они также производятся с использованием определенной закваски и культуры ПКБ. В отличие от Emmentaler PDO, эти сыры являются полутвердыми сырами, изготовленными из пастеризованного коровьего молока. Они характеризуются мягкой и гладкой текстурой с крупными круглыми отверстиями и мягким ореховым вкусом. Время их созревания обычно очень короткое по сравнению с Emmentaler PDO; например, Maasdam готов к употреблению уже через 4 недели созревания. Поскольку технология производства основана на скрещивании Гауды и Эмменталя, эту группу иногда называют "Гуталер" («Goutaler») (Abrahamsen et al., 2006). В отличие от сыров швейцарского типа с защищенным наименованием места происхождения, эти сыры часто разрабатываются и производятся в рамках одной конкретной компании и представляют собой торговую марку, как, например, Jarlsberg. Поэтому трудно найти технологическую информацию о переработке и созревании таких сыров швейцарского типа.
D. Сыры со спонтанным пропионовокислым брожением
Comté PDO (Комте) - французский сорт твердого сыра, который может подвергаться спонтанному пропионовокислому брожению, причем ПКБ происходят из сырого молока. Этот сорт сыра не обязательно похож на швейцарские сыры, но может иметь глазки или трещины, а также "фруктовый" привкус. В настоящее время большинство сыров Комте PDO не подвергаются пропионовокислому брожению, однако оно допускается. Кроме того, полумягкий итальянский сыр Fontina PDO (Фонтина) производится из сырого молока и часто подвергается спонтанной ферментации ПКБ. Сыр Фонтина обычно испещрен мелкими дырочками; вкус мягкий, фруктовый, с возрастом приобретает сильные ореховые и грибные нотки (рис. 35.1).
E. Различие между "швейцарским сыром" и "сыром швейцарского типа"
Применение общих терминов "сыр швейцарского типа" и "швейцарский сыр" часто приводит к путанице. Например, швейцарский сыр "Грюйер" PDO не подвергается пропионовокислому брожению, однако часто ошибочно считается сыром швейцарского типа. Напротив, "Грюйер французский" PGI - это твердый сыр швейцарского типа, который подвергается пропионовокислому брожению (рис. 35.1). Аналогичным образом, различные полутвердые сорта сыра из Швейцарии, такие как Аппенцеллер, Тильзит и Раклетт, а также голландский сорт Гауда, часто рассматриваются как сыры швейцарского типа из-за их типичной открытой текстуры с небольшими круглыми отверстиями или из-за страны происхождения. Однако CO2 для образования отверстий поступает не из ПКБ, а из других источников - в большинстве случаев из гетероферментативных молочнокислых бактерий (Martley and Crow, 1996). Поэтому рост ПКБ в вышеупомянутых сортах сыра можно считать дефектным. В США швейцарский сыр по-прежнему является синонимом эмментальского сыра (FDA, 2014). В данной главе термин "сыр швейцарского типа" используется для сыров с характерным пропионовокислым брожением, получаемым путем добавления ПКБ в сырное молоко.
ПРОПИОНОВОКИСЛОЕ БРОЖЕНИЕ И ВЗАИМОДЕЙСТВИЯ МЕЖДУ МИКРООРГАНИЗМАМИ В СЫРЕ

Пропионовокислое брожение
Пропионовокислое брожение — это превращение сахара или молочной кислоты и ее солей в пропионовую и уксусную кислоты с выделением углекислого газа и воды. Некоторые пропионовокислые бактерии образуют, кроме того, небольшое количество других кислот (муравьиной, янтарной, изовалериановой). При пропионовокислом брожении превращение глюкозы до пировиноградной кислоты протекает также по гликолитическому пути. В дальнейшем пировиноградная кислота, претерпевая ряд превращений, восстанавливается в пропионовую.
Среди четырех "молочных ПКБ" (Propionibacterium freudenreichii, Propionibacterium acidipropionici, Propionibacterium thoenii и Propionibacterium jensenii) при производстве сыров с пропионовокислым брожением для получения характерных глазков и орехового привкуса предпочтительно используются штаммы вида P. freudenreichii. В традиционном Эмменталере PDO инокуляционная доза ПКБ составляет 103-104 КОЕ/мл сырого молока (Turgay et al., 2011). В пастеризованных сырах швейцарского типа ПКБ добавляют в сырное молоко в начале процесса производства, чтобы обеспечить их присутствие в количестве около 103 КОЕ/г сыра после производства (Beresford et al., 2001). Они хорошо развиваются в сыре, начиная с низких количеств, но не делают этого в молоке (Piveteau et al., 2000). Чтобы ускорить и контролировать рост ПКБ в сырах швейцарского типа, процесс созревания делится на десять периодов, в которых чередуются условия. Например, при производстве Эмменталера PDO сыры сначала хранятся в холодном помещении при температуре (12°C) в течение 10 дней, затем в теплом помещении (при 22°C, 80% относительной влажности, RH) в течение 60 дней, и, наконец, дозревают в холодном помещении (12°C, 70% RH). Несмотря на значительные различия в процедурах созревания сыров швейцарского типа, в сырах, готовых к употреблению, ПКБ присутствуют на уровне примерно 108-109 КОЕ/г.
На рис. 35.2 показано развитие ПКБ в сыре Эмменталер PDO в процессе производства и созревания. Сыры были произведены с использованием культур ПКБ с различной специфической аспартазной активностью: сыр А - с низкой, сыр В - с высокой и сыр С - со средней специфической аспартазной активностью.
 во время традиционного созревания Эмменталера PDO, произведенного с использованием культур ПКБ с различной специфической аспартазной активностью")
Рисунок 35.2 Развитие пропионовокислых бактерий (ПКБ КОЕ/г) во время традиционного созревания Эмменталера PDO, произведенного с использованием культур ПКБ с различной специфической аспартазной активностью: сыры A, B и C с низкой, высокой и средней специфической аспартазной активностью, соответственно. (Перепечатано из Turgay, et al., 2011. Biodiversity, dynamics, and characteristics of Propionibacterium freudenreichii in Swiss Emmentaler PDO cheese. Dairy Sci.Technol. 91, 471–489.)
Пропионибактерии грамположительны, малоподвижны, неспорообразующи и выглядят под микроскопом как короткие палочки, морфология которых заметно зависит от условий и фазы культуры (рис. 35.3). Они растут только при низких концентрациях кислорода (от анаэробных до аэротолерантных), чувствительны к соли и имеют оптимальный рост при рН между 6 и 7 с максимумом 8,5 и минимумом 4,6 (речь о росте, а не о выживании – ред.) (Langsrud and Reinbold, 1973).

Рисунок 35.3 Сканирующая электронная микрофотография лиофилизированной культуры (слева) и световая микрофотография (справа) жидкой культуры Propionibacterium freudenreichii (Agroscope, Bern, Switzerland).
Чувствительность к соли зависит от штамма ПКБ, а также от изменения рН: они переносят 6% соли при рН 7, но только 3% при рН 5,2. Richoux et al. (1998) сообщили, что увеличение содержания соли во влаге с 1 до 3 % заметно снижает содержание летучих жирных кислот и выработку CO2 в сыре швейцарского типа. Низкая концентрация NaCl в сырах швейцарского типа благоприятствует росту ПКБ; сыр Эмменталер PDO обычно содержит 0,5% (о массе), а французский Эмменталь - 0,4-0,7%. Для производства сыров швейцарского типа со средним или высоким содержанием соли (Grevé 1,1-1,4 %, Maasdam 1,5 % и Fol-Epi 1,6 %) необходимо использовать адаптированные культуры ПКБ. Оптимальная температура роста колеблется от 25 до 35°C, но рост также происходит при 14°C и ниже и зависит от штамма (Fröhlich-Wyder and Eugster-Meier, 2005; Ojala et al., 2017; Park et al., 1967; Turgay et al., 2011). Пропионибактерии термотолерантны и могут выдерживать температуру 50-55°C в течение 30 минут (Thierry et al., 2011). Кроме того, выделение ПКБ из сырого молока с помощью культурально-зависимых методов выявило доминирование вида P. freudenreichii, что свидетельствует о том, что этот вид является наиболее устойчивым к холодовому и тепловому стрессу среди молочных ПКБ.
Производство пропионовой кислоты в ПКБ функционирует как циклический процесс, в котором одновременно участвуют несколько взаимосвязанных путей (рис. 35.4 по Crow et al., 1988; Thierry et al., 2011). Лактат используется в качестве источника энергии, а аспартат - в качестве акцептора электронов, причем оба эти вещества имеются в сыре (Brendehaug and Langsrud, 1985; Crow, 1986; Crow and Turner, 1986). Основными продуктами ферментации являются пропионовая, уксусная, янтарная кислоты и CO2 (реакции D и E на рис. 35.4). В основном лактат, а также аланин и серин превращаются в пируват с образованием АТФ и восстановленных коферментов (Ojala et al., 2017; Thierry et al., 2011). Пируват либо окисляется до ацетата и CO2 (реакция B), либо восстанавливается до пропионата по пути Вуда-Веркмана (реакция C). Ферменты пути Вуда-Веркмана были идентифицированы в геноме P. freudenreichii (Falentin et al., 2010; Ojala et al., 2017) и P. acidipropionici (Parizzi et al., 2012). Пропионибактерии регулируют пропорции пирувата, который либо окисляется, либо восстанавливается в зависимости от используемого субстрата, условий окружающей среды и характеристик штамма (Ojala et al., 2017; Thierry et al., 2011).
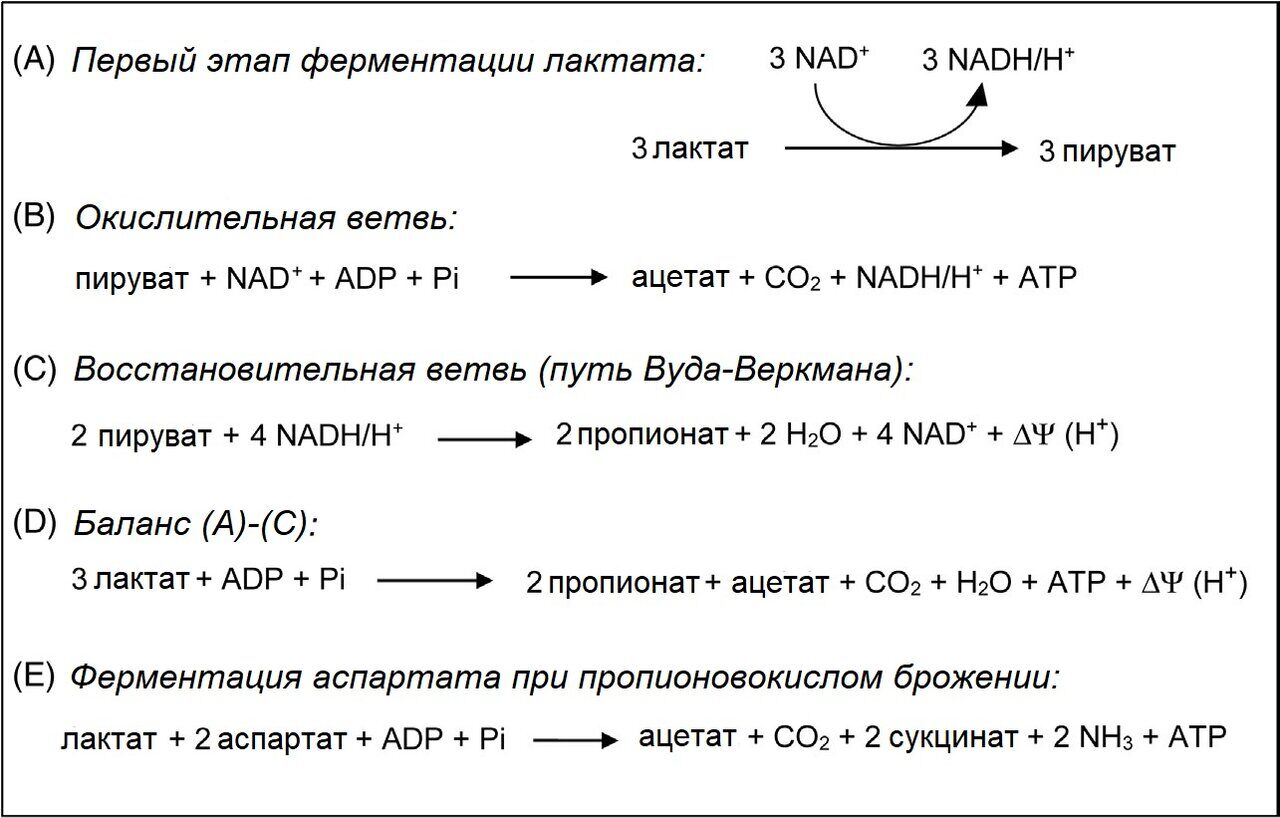
Рисунок 35.4 Метаболические пути утилизации лактата пропионовокислыми бактериями. (Адаптировано из Crow, V.L., Turner, K.W., 1986. The effect of succinate production on other fermentation products in Swiss-type cheese. New Zeal. J. Dairy Sci. 21, 217–227 и Thierry, et al., 2011. New insights into physiology and metabolism of Propionibacterium freudenreichii. Int.J. Food Microbiol. 149, 19–27.)
В присутствии аспартата метаболизм лактата соединяется с метаболизмом аспартата, который дезаминируется до фумарата и далее восстанавливается до сукцината (рис. 35.4, реакция Е). Пируват из лактата в основном окисляется до ацетата и CO2. Следовательно, для поддержания окислительно-восстановительного баланса клетки больше лактата ферментируется до ацетата и CO2, чем до пропионата. Штаммы P. freudenreichii могут сильно отличаться по своей специфической аспартазной активности (Blasco et al., 2011; Richoux and Kerjean, 1995; Turgay et al., 2011). Пропионибактерии с низкой удельной аспартазной активностью способны метаболизировать in vitro не более 100 нмоль аспартата/мин (Fröhlich-Wyder et al., 2002). Штаммы с высокой специфической аспартазной активностью способны метаболизировать in vitro до 800 нмоль аспартата/мин. Следовательно, обычно весь доступный аспартат метаболизируется до сукцината (рис. 35.5).
 и аспарагина (Asn) в 6-месячном выдержанном сыре Эмменталер PDO")
Рисунок 35.5 Линейная регрессия с 95% доверительным интервалом для сукцината и суммы аспартата (Asp) и аспарагина (Asn) в 6-месячном выдержанном сыре Эмменталер PDO (▼, ПКБ с высокой специфической аспартазной активностью; ▲, ПКБ с низкой специфической аспартазной активностью) (Перепечатано из Wyder, M.T., Bosset, J.O., Casey, M.G., Isolini, D. and Sollberger, H., 2001. Influence of two different propionibacterial cultures on the characteristics of Swiss-type cheese with regard to aspartate metabolism. Milk Sci. Int. 56, 78-81).
Высокая удельная активность аспартазы обычно сочетается с более высокой скоростью роста ПКБ, что приводит к более высоким показателям и концентрациям пропионата, ацетата и CO2 (таблица 35.1, рис. 35.2 и 35.6). Аналогичным образом, Piveteau et al. (1995) показали, что скорость роста и выход ПКБ в сыворотке можно повысить путем добавления аспартата.
На открытость сыров швейцарского типа большое влияние оказывает специфическая аспартазная активность применяемых ПКБ и ПКБ-флоры, уже присутствующей в сыром молоке, как показано в работе Turgay et al. (2011). В результате повышенного выделения CO2 количество и размер глазков увеличиваются, а высота буханок больше при использовании культур ПКБ с высокой специфической аспартазной активностью (рис. 35.7). Время созревания сыров в теплом помещении может сократиться до 10 дней (Fröhlich-Wyder et al., 2002). Однако в результате продолжающегося пропионовокислого брожения, как видно на рис. 35.6, такие сыры более склонны к позднему брожению, что нежелательно, если сыры созревают до 12 месяцев или дольше (табл. 35.1).
Использование культур ПКБ с высокой специфической аспартазной активностью может быть выгодно в сырах швейцарского типа с более коротким периодом созревания, поскольку они способствуют более выраженному вкусу (Fröhlich-Wyder et al., 2002; Wyder et al., 2001). Основная причина, по-видимому, заключается в более высоком содержании летучих карбоновых кислот, выделяющихся в процессе ферментации (рис. 35.6), и свободных жирных кислот, n-масляной и n-капроновой, которые высвобождаются в результате липолитической активности ПКБ (табл. 35.1). Это комбинированный эффект метаболизма аспартата и увеличения количества пропионовокислых бактерий. Таким образом, для применения культур ПКБ в производстве сыра необходимо учитывать способность утилизировать аспартат.

Рисунок 35.6 Разложение лактата (-▲-) и образование пропионата (- - -■- - -) и ацетата (⋯♦⋯) в сыре Эмменталер PDO, произведенном с использованием культур пропионибактерий с высокой специфической аспартазной активностью (A) и низкой специфической аспартазной активностью (B). (Адаптировано из Wyder, M.T., Bosset, J.O., Casey, M.G., Isolini, D., Sollberger, H., 2001. Influence of two different propionibacterial cultures on the characteristics of Swiss-type cheese with regard to aspartate metabolism. Milk Sci. Int. 56, 78-81).
 и высокой (сыр № 11) специфической аспартазной активностью")
Рисунок 35.7 Эмменталер PDO в 12 месяцев, изготовленный с использованием культур ПКБ с низкой (сыр № 9) и высокой (сыр № 11) специфической аспартазной активностью (Agroscope, Bern, Switzerland) (см. раздел " Color Plate ").
Таблица 35.1 Средние значения метаболитов, свободных аминокислот и количества пропионовокислых бактерий в сыре Эмменталер PDO (6 и 12 месяцев), изготовленном с использованием культур ПКБ с низкой и высокой удельной аспартазной активностью.
|
Параметр
|
Эмменталер PDO 6 месяцев
|
Эмменталер PDO 12 месяцев
|
||||
|
Низкое
значение
(N = 10)
|
Высокое значение
(N = 8)
|
t-Тест
|
Низкое
значение
(N = 10)
|
Высокое значение
(N = 8)
|
t-Тест
|
|
| Лактатa | 57.4 ± 10.5 | 45.3 ± 17.4 | ns | 47.0 ± 8.5 | 11.3 ± 6.7 | *** |
| Знгачение pH | 5.75 ± 0.02 | 5.79 ± 0.02 | ** | 5.63 ± 0.06 | 5.73 ± 0.02 | *** |
| Свободные SCAa | 114.4 ± 5.2 | 126.0 ± 5.2 | ns | 117.4 ± 5.9 | 148.1 ± 5.0 | *** |
| Ацетатa | 48.4 ± 1.3 | 53.1 ± 5.1 | * | 47.6 ± 0.6 | 58.7 ± 1.7 | *** |
| Пропионатa | 60.1 ± 4.4 | 67.1 ± 10.2 | ns | 63.2 ± 4.2 | 83.6 ± 3.6 | *** |
| n-Бутиратa | 1.1 ± 0.2 | 1.2 ± 0.1 | ns | 1.7 ± 0.9 | 1.7 ± 0.1 | ns |
| n-Капронатa | 0.4 ± 0.1 | 0.5 ± 0.1 | * | 0.5 ± 0.1 | 0.7 ± 0.1 | ** |
| Сукцинатa | 4.0 ± 0.6 | 11.9 ± 1.7 | *** | 5.1 ± 2.8 | 17.7 ± 2.5 | *** |
| CO2a | 27.6 ± 1.6 | 33.6 ± 2.0 | *** | nd | nd | |
| Пропионибактерииb | nd | nd | 6.7 ± 0.9 | 8.4 ± 0.3 | *** | |
| Свободные аминокислотыa | 169.0 ± 23.7 | 165.5 ± 30.2 | ns | 266.9 ± 34.5 | 246.9 ± 23.0 | ns |
| Аспартатa | 2.2 ± 0.9 | 0.0 ± 0.0 | *** | 4.8 ± 0.6 | 0.6 ± 0.1 | *** |
| Аспарагинa | 2.9 ± 1.1 | 0.1 ± 0.2 | *** | 1.9 ± 0.5 | 0.1 ± 0.2 | *** |
nd, не определено; ns, незначительно; * P < 0.05; ** P < 0.01; *** P < 0.001; SCA, короткоцепочечные кислоты; a ммоль/кг; b log КОЕ/г. (Перепечатано из Wyder, M.T., Bosset, J.O., Casey, M.G., Isolini, D. and Sollberger, H., 2001. Влияние двух различных культур пропионибактерий на характеристики сыра швейцарского типа с точки зрения метаболизма аспартата. Milk Sci. Int. 56, 78-81).
Взаимодействие пропионовокислых бактерий (ПКБ) с молочнокислыми бактериями (МКБ)
В сырах швейцарского типа комменсализм между МКБ и ПКБ существует в источнике углерода, где МКБ ферментируют лактозу до молочной кислоты, которая впоследствии используется ПКБ (Smid and Lacroix, 2013). Однако не только наличие или отсутствие молочной кислоты, но и скорость производства кислоты может влиять на рост ПКБ; медленное производство кислоты МКБ стимулирует рост ПКБ, в то время как быстрое производство кислоты оказывает ингибирующее действие (Perez Chaia et al., 1987; Piveteau et al., 1995). Помимо молочнокислого брожения, на пропионовокислое брожение существенное влияние оказывают и другие факторы, такие как сезон производства молока (Daly et al., 2010), температура, значение pH, содержание солей и протеолиз.
Мезофильные закваски
Для производства различных полутвердых сыров швейцарского типа, таких как Ярлсберг, Маасдам или Гревé, используются мезофильные молочнокислые бактерии, которые традиционно присутствуют в заквасках для производства сыров голландского типа, таких как Гауда. Эти заквасочные культуры содержат молочнокислые кокки, а именно Leuconostoc spp. и Lactococcus lactis ssp. lactis biovar diacetylactis (Miks-Krajnik et al., 2013). Мезофильные закваски, используемые для производства Ярлсберга или Гревé, включают L. lactis ssp. lactis, L. lactis ssp. cremoris, L. lactis ssp. lactis biovar diacetylactis (L. diacetylactis; цитрат-положительный) и различные подвиды Leuc. mesenteroides (Ardö and Varming, 2010). Лактококки - это гомоферментативные микроаэрофильные грамположительные бактерии, которые растут при температуре 10°C, но не при 45°C, и производят L(+)-молочную кислоту из глюкозы (Samarzija et al., 2001). По сравнению со штаммами L. lactis ssp. cremoris, штаммы L. lactis ssp. lactis обычно менее чувствительны к температуре и способствуют быстрому и воспроизводимому подкислению сыра (Ardö and Varming, 2010). Leuconostoc spp. гетероферментативно метаболизируют лактозу до D(−)-лактата, этанола и CO2 и растут при более низком pH, чем лактококки, что дает им преимущество в конце процесса подкисления сыра.
До периода размещения в теплом помещении L. diacetylactis и Leuconostoc spp. преобразуют большую часть лимонной кислоты в сыре в уксусную кислоту, диацетил, ацетоин, 2,3-бутандиол и CO2 и тем самым инициируют образование глазков (Ardö and Varming, 2010; Rehn et al., 2011). В зависимости от условий обработки и созревания сыра, любая из этих двух цитрат-положительных групп может доминировать (Ardö and Varming, 2010).
Термофильные закваски
При производстве твердых сортов сыра термофильные молочнокислые бактерии обычно используются в заквасках, часто в виде смешанных культур лактобацилл (Lactobacillus helveticus, Lactobacillus delbrueckii ssp. lactis) и стрептококков (Streptococcus salivarius ssp. thermophilus). Они гарантируют гомоферментативный катаболизм лактозы до >90% лактата. Стрептококки производят только L(+)-молочную кислоту, тогда как Lb. delbrueckii ssp. lactis полностью превращает лактозу в D(-)-лактат. Lb. helveticus продуцирует оба изомера. Лактоза полностью гидролизуется в течение 4-6 ч после формования сырной массы, а молочнокислое брожение завершается через 24 ч. Галактоза, образующаяся при гидролизе лактозы, не используется стрептококками, а далее метаболизируется лактобациллами. Чтобы избежать нежелательного брожения во время созревания, в конце процесса молочнокислого брожения не должно оставаться остатков галактозы.
Несколько исследований показали, что термофильные молочнокислые бактерии, в частности Lb. delbrueckii и Lb. helveticus, способны стимулировать рост пропионибактерий (Chamba, 2000; Kerjean et al., 2000; Perez Chaia et al., 1987; Piveteau et al., 1995). Было показано, что Lb. helveticus лизируется на ранних стадиях созревания сыра, в результате чего цитоплазматические ферменты, такие как пептидазы, высвобождаются в матрицу сыра (Deutsch et al., 2002). Baer и Ryba (1999) обнаружили, что ПКБ предпочитают свободные аминокислоты пептидам, причем аспартат является аминокислотой, которая наиболее легко метаболизируется (Piveteau et al., 1995). Напротив, Piveteau et al. (2002) показали, что пептиды, продуцируемые Lb. helveticus, оказывают большее стимулирующее действие на рост ПКБ, чем аминокислоты. Те же авторы привели доказательства наличия в молоке ингибирующего вещества, которое термостабильно и имеет низкую молекулярную массу (Piveteau et al., 2000). Оно удаляется штаммами Lb. helveticus в результате протеолиза, но не штаммами Lb. delbrueckii или Lb. lactis. Следовательно, стимуляция роста пропионовокислых бактерий может быть результатом протеолитической активности лактобацилл, высвобождающих пептиды и свободные аминокислоты, и/или удаления ингибирующего вещества под действием Lb. helveticus (Kerjean et al., 2000). Несколько десятилетий назад, в том числе при производстве Эмменталера PDO, Lb. helveticus был основным компонентом заквасочных культур. Из-за интенсивной протеолитической активности и стимулирующего влияния на рост пропионовокислых бактерий он часто ассоциировался с поздней ферментацией. По этой причине он был успешно заменен на Lb. delbrueckii ssp. lactis.
Нестартерные молочнокислые бактерии
Факультативно гетероферментативные нестартерные лактобациллы (FHL или ФГЛ) либо присутствуют в качестве местной микрофлоры в сыром молоке сыров швейцарского типа, либо целенаправленно используются при производстве сыров швейцарского типа (Bouton et al., 2009; Fröhlich-Wyder et al., 2002). Они ферментируют гексозы почти исключительно до молочной кислоты. В эту группу микроорганизмов входят, в частности, Lactobacillus casei и Lactobacillus rhamnosus. Во время созревания сыра они растут, утилизируя цитрат, который содержится в недозрелом сыре, и тем самым могут инициировать образование глазков (O'Sullivan et al., 2016; Rehn et al., 2011). Начиная с 9 ммоль/кг цитрата в сыре, местные ФГЛ используют примерно 3 ммоль, а те, что добавлены в качестве дополнительной культуры, метаболизируют весь доступный цитрат до муравьиной кислоты, уксусной кислоты и CO2 (Fröhlich-Wyder et al., 2002; Martley and Crow, 1996). ФГЛ начинают рост в начале созревания и достигают примерно 108 КОЕ/г (Thierry et al., 1998).
При производстве финского эмменталя ФГЛ используются в качестве защитных штаммов, особенно против клостридий (Rodriguez et al., 2011). В сырах, таких как Комте, изготовленных из сырого молока, они необходимы для улучшения вкуса (Bouton et al., 2009), а также могут применяться для модификации определенных вкусовых качеств сыра (Kocaoglu-Vurma et al., 2008). Более того, ФГЛ используются в Швейцарии для замедления пропионовокислого брожения (Sollberger and Wyder, 2000). Jimeno et al. (1995) обнаружили ингибирование роста ПКБ в сыре до 80 % по сравнению с контролем без ФХЛ (L. casei и L. rhamnosus). Следовательно, образуется меньше пропионовой кислоты (Martley and Crow, 1996). Способность ФГЛ метаболизировать цитрат играет важную роль, поскольку цитрат-негативные мутанты, как было показано, ингибируют ПКБ гораздо меньше, чем соответствующие цитрат-позитивные штаммы (Weinrichter et al., 2004). Lactobacillus rhamnosus также продуцирует небольшое, но заметное количество диацетила, который оказывает летальное действие на ПКБ (Baer and Ryba, 1999). Ацетат и формиат, по-видимому, также оказывают ингибирующее действие на рост ПКБ. Кроме того, метаболизм цитрата, который происходит перед пропионовокислым брожением, приводит к высвобождению закомплексованной меди. Относительная концентрация цитрата и меди играет важную роль в наблюдаемом ингибировании (Perez Chaia et al., 1987). Однако механизм ингибирования пока окончательно не выяснен.
Биоразнообразие и динамика ПКБ в сыре
Пропионибактерии естественным образом встречаются в рубце и кишечнике жвачных животных, в почве и силосе, поэтому они также присутствуют в сыром молоке в небольшом количестве. Штаммовое разнообразие естественной пропионибактериальной флоры велико, и на него, к счастью, не повлияло широкое использование коммерчески доступных культур (Fessler, 1997). Turgay et al. (2011) оценили распространенность штаммов ПКБ дикого типа в сыре Эмменталер PDO и их влияние на качество сыра. В результате воздействия высокой температуры ошпаривания (2-го нагревания) из творога или созревшего сыра не удалось выделить ни одного вида, кроме P. freudenreichii, хотя в сыром молоке, использованном для производства сыра, изначально присутствовали другие виды молочных ПКБ. Исследование также показало, что в процессе созревания сыра происходят важные изменения в составе флоры P. freudenreichii.
, полученного с добавлением штаммов ПКБ с низкой специфической аспартазной активностью, после сроков созревания 2 и 8 месяцев")
Рисунок 35.8 Сравнение состава ПКБ-флоры сыра А (Эмменталер PDO), полученного с добавлением штаммов ПКБ с низкой специфической аспартазной активностью, после сроков созревания 2 и 8 месяцев. Размер кружков соответствует процентному содержанию штаммов индивидуальной культуры (А) и дикого типа (WT); положение кружков показывает потенциал роста при 11°С и специфическую активность аспартазы штаммов. (Генотип А1 представляет два штамма. Два кружка одинакового размера обозначают общий процент генотипа А1). (Перепечатано из Turgay, et al., 2011. Biodiversity, dynamics, and characteristics of Propionibacterium freudenreichii in Swiss Emmentaler PDO cheese. Dairy Sci.Technol. 91, 471–489.)
Хранение в теплом помещении (22°C) в целом благоприятствовало росту штаммов дикого типа P. freudenreichii. Однако при последующем хранении в холоде (11 °C) наблюдались заметные различия в количестве генотипов и соотношении штаммов стартового и дикого типа. В Эмменталере PDO, изготовленном с добавлением двух штаммов ПКБ с низкой специфической аспартазной активностью (сыр А), доля штаммов дикого типа достигла 81 % через 8 месяцев созревания (рис. 35.8). Этот пример наглядно иллюстрирует, что созревание сыра - это непрерывный процесс отбора, который приводит к резким изменениям в штаммовом составе даже в популяции бактерий одного вида. Сильный рост более адаптированных штаммов дикого типа P. freudenreichii в этом сыре был в основном связан с сильным ингибированием добавленной культуры ПКБ со стороны ФГЛ и высокой специфической аспартазной активностью растущих штаммов дикого типа, что указывает на то, что эти два свойства являются ключевыми факторами в контроле пропионовокислого брожения. Важно отметить, что ПКБ с высокой специфической аспартазной активностью в меньшей степени ингибируются ФГЛ. При производстве Эмменталера PDO совместное использование культуры ПКБ с низкой удельной аспартазной активностью и добавочной культуры Lb. casei требует длительного пребывания сыров в теплом помещении, в то время как использование культуры ПКБ с высокой удельной аспартазной активностью и отсутствие добавок ФГЛ приводит к сокращению времени пребывания (Fröhlich-Wyder et al., 2002). Можно производить сыры швейцарского типа с глазками, сделанными по мерке: большие глазки на относительно ранней стадии достижимы при совместном использовании Lb. helveticus и высокоспецифичной аспартазоположительной культуры ПКБ. Глазки меньшего размера получаются при использовании вспомогательных культур ФГЛ вместе со слабоспецифичной аспартаза-положительной культурой ПКБ. Однако конструкция таких культуральных систем для производства сыров швейцарского типа и наличие штаммов дикого типа влияют не только на размер глазков, но и на способность к созреванию сыров швейцарского типа, изготовленных из сырого молока. Для производства полутвердых сыров швейцарского типа из пастеризованного молока, которые поступают в продажу после короткого периода созревания, добавление штаммов P. freudenreichii с высокой специфической аспартазной активностью и отсутствие добавок ФГЛ может дать сыры с оптимальными глазками и вкусообразованием. В отличие от этого, такая система культивирования не подходит для производства традиционного Эмменталера PDO, рассчитанного на более длительный период созревания, поскольку позволяет получать сыры, склонные к поздней ферментации. Важно также отметить, что совместное использование культур ПКБ с высокой и низкой удельной аспартазной активностью не дает сыров с промежуточными свойствами с точки зрения глазков, формирования вкуса и способности к созреванию. Данные, представленные на рис. 35.8, и практический опыт сыроделов показывают, что штаммы P. freudenreichii с высокой удельной аспартазной активностью будут доминировать в таких культуральных системах, даже если они добавляются в сырное молоко в меньшем количестве.
ТЕХНОЛОГИЯ

Обработка молока
Молоко, используемое для производства сыра, должно содержать как можно меньше бактерий, чтобы добавленные заквасочные культуры могли оказать оптимальное воздействие. Если используется сырое молоко, бактериологические требования особенно строги. Разумеется, микробное и гигиеническое состояние фермерского молока зависит также от продолжительности и температуры хранения; необходимо избегать вторичного загрязнения до, во время или после переработки. В Швейцарии сыр Эмменталер PDO должен производиться из сырого молока коров, не получающих силосных кормов, а молоко должно быть превращено в сыр в течение 24 часов после дойки (Anonymous, 2012). Однако сыры швейцарского типа, такие как Маасдам, Ярлсберг, Греве или Эмменталь, производятся из пастеризованного молока, которое подвергается бактофугированию, чтобы удалить споры клостридий и тем самым предотвратить маслянокислое брожение (Rehn et al., 2011; Rodriguez et al., 2011). Другие меры по борьбе с клостридиями включают добавление консервантов, таких как нитраты (Abrahamsen et al., 2006; Reinbold, 1972; Scott, 1998), лизоцим и низин, или использование антиклостридиальных вспомогательных культур (Rodriguez et al., 2011).
Производство
Вся технологическая последовательность операций направлена на создание оптимальных условий для пропионовокислого брожения и образования глазков. Основополагающим этапом является добавление воды в молоко и/или сырную массу (12%-20% для Эмменталера PDO; ∼25%-30% для Ярлсберга или финского Эмменталя) для снижения концентрации лактозы. Это приводит к относительно высокому значению pH после молочнокислого брожения (pH 5,20-5,35), что, соответственно, ускоряет пропионовокислое брожение. Кроме того, ограничение выработки молочной кислоты снижает потери кальция в сыворотке. Высокое содержание кальция в твороге (сырной массе) приводит к мягкой и эластичной текстуре, что очень важно для регулярного формирования глазков.
Для большинства полутвердых сыров швейцарского типа температура варки или ошпаривания (2-го нагревания) составляет около 40°C. Ярлсберг после нарезки нагревают до 36-39°C (Abrahamsen et al., 2006), а Греве - до 40°C (Rehn et al., 2011). В отличие от этого, при производстве твердого Эмменталя (например, финского, французского или швейцарского) температура ошпаривания составляет 51-54°C (Anonymous, 2012; Rodriguez et al., 2011). Во время прессования твердых сыров большого размера температура в сырной массе остается на уровне 50°C в течение многих часов. При такой температуре из творога вытекает больше сыворотки и инактивируется большинство нежелательных микроорганизмов. Это важно для обеспечения гигиенической безопасности, особенно сыров из сырого молока. Твердые сыры обычно являются долгосозревающими сырами, поэтому высокая температура ошпаривания и прессования важна для того, чтобы избежать слишком интенсивного протеолиза (пептидолиза), который приведет к "укороченной" или более рассыпчатой текстуре и увеличит риск появления расслоений и трещин во время созревания. Высокая температура приготовления также приводит к полной инактивации химозина. Однако температура выше 54°C сильно ухудшает рост ПКБ.
Традиционные сыры швейцарского типа, такие как Эмменталер PDO, производятся в чанах из меди (Cu) и прессуются в круглых формах (рис. 35.9). В Финляндии производство сыра Эмменталь осуществляется в чанах из нержавеющей стали, а медь добавляется в виде раствора сульфата меди на уровне, обеспечивающем концентрацию меди в конечном сыре, близкую к концентрации меди в традиционном сыре Эмменталер PDO (Rodriguez et al., 2011). Содержание меди в сыре Эмменталер PDO обычно составляет от 7,6 до 16,5 мг/кг (Sieber et al., 2006); самый высокий допустимый уровень в финском эмментальском сыре - 15,0 мг/кг (Rodriguez and Alatossava, 2008). Содержание Cu не должно быть слишком высоким, поскольку Cu препятствует образованию молочной и пропионовой кислот. С другой стороны, Cu может замедлить пропионовокислое брожение и ингибировать прорастание спор C. tyrobutyricum, что снижает риск возникновения позднего пузырения (Rodriguez and Alatossava, 2010). Кроме того, Cu образует комплексы с сернистыми соединениями, образующимися при катаболизме аминокислот, и таким образом оказывает положительное влияние на вкус и аромат сыра (Sieber et al., 2006). Однако для производства многих других сыров швейцарского типа используются сырные чаны из нержавеющей стали без добавления Cu.

Рис. 35.9. Нарезка сырной массы в медной чане, заполнение сырной массы в формы для сыра Эмменталь и засолка Эмменталера PDO. (Из Википедии и Дорф-Часи, Хергисвиль, Швейцария.)
Просолку проводят после прессования, после достижения значения рН 5,20–5,35. Процесс засолки приводит к получению твердой и сухой кожуры. Поглощение и распространение соли в сырах швейцарского типа сильно зависит от размера и формы буханок. Хотя колеса Эмменталера PDO маринуются в течение 2 дней (рис. 35.9), поглощение соли остается очень скромным по сравнению с большим весом сыров (75–120 кг). В некоторых сырах швейцарского типа соль добавляют в творожно-сывороточную смесь во время производства (Abrahamsen et al., 2006; Reinbold, 1972). В результате таких различий содержание соли в сырах швейцарского типа значительно варьируется (например, Эмменталер PDO ∼0,5%, Ярлсберг ∼1,25%, Фоль-Эпи ∼1,6%). В настоящее время сыры швейцарского типа производятся во многих странах по технологиям, отличным от оригинальных. Что касается технологических аспектов, сыры швейцарского типа всегда варятся. С другой стороны, обработка молока, степень механизации, использование добавок, вес и форма, средний состав (твердые или полутвердые сыры, оба с разным содержанием жира) и время созревания часто сильно различаются.
Условия созревания

Сухое созревание применяется в основном при созревании традиционных сыров швейцарского типа. Однако большинство сыров швейцарского типа созревают в пластиковой пленке или воске, как, например, Ярлсберг, Греве и Маасдам. Созревание швейцарских сыров состоит из трех основных периодов: период сушки и расплавления творога, период выдержки в теплом помещении и период окончательного созревания. Период сушки обычно проводится при температуре 4-12°C и длится до 2 недель. В теплое время года сыры помещают в помещение при температуре 17-25°C на 3-8 недель. Окончательное созревание происходит в холодном помещении при температуре 4-12°C, чтобы замедлить выработку CO2 (Chen et al., 2009; Ji et al., 2004; Rehn et al., 2011; Sheehan et al., 2008). Продолжительность периода выдержки в теплом помещении и окончательного созревания в значительной степени зависят от сорта и, следовательно, могут значительно варьироваться. Рис. 35.10 иллюстрирует влияние повышения температуры в теплом помещении (DT на 3°C между вариантами) на развитие объема глазков.
При производстве традиционных швейцарских сыров, таких как Emmentaler PDO, продолжительность выдержки в теплом помещении оценивается путем прослушивания звука при постукивании по поверхности сыра специальным молотком. Во время формирования глазка звук постукивания меняется, и опытный сыровар знает, когда сыры готовы к последнему периоду созревания в холодильной камере.
, T(0) и T(1) с перепадом 3°C между каждым уровнем")
Рис. 35.10 Развитие объема глазка в сыре во время созревания в теплом помещении при температурах T(-1), T(0) и T(1) с перепадом 3°C между каждым уровнем. Объем глазка был рассчитан с использованием "пикселя" в качестве меры длины для оценки радиуса отдельных глазков. (Перепечатано из Kraggerud, H., Wold, J.P., Hoy, M. and Abrahamsen, R.K., 2009. X-ray images for the control of eye formation in cheese. Int. J. Dairy Technol. 62, 147-153.)
Относительная влажность воздуха во время созревания Эмменталера PDO довольно низкая (70-80 %). Это, наряду с рассолом, приводит к образованию твердой и сухой кожуры, которая уменьшает потерю CO2 во время пропионовокислого брожения, тем самым способствуя образованию глазков. Упругая кожура также обеспечивает прочную форму во время созревания в теплом помещении. Кроме того, низкая влажность ускоряет так называемое "потоотделение" сыров (выделение жира) и уменьшает рост плесени на поверхности сыров и, следовательно, время, необходимое для ручной очистки. Некоторые сыры швейцарского типа (например, Gruyère français PGI) созревают во влажных подвалах (>90% RH; относительной влажности) при температуре от 12 до 18°C и регулярно обрабатываются водой, в результате чего на их поверхности развивается мазковая флора. Кроме того, отборные сорта Эмменталера PDO переносятся во влажные подвалы (90% RH) в возрасте 4 месяцев, инокулируются на поверхности Penicillium sp. и дозревают еще 6-8 месяцев. На поверхности образуется темный налет, уменьшается потеря воды во время созревания, а значит, протеолиз и формирование вкуса протекают с большей скоростью, чем в сырах сухого созревания.
Промышленно производимые сыры швейцарского типа обычно покрывают воском или заворачивают в пластиковую пленку. Это существенно снижает затраты на обработку сыра во время созревания, минимизирует потерю влаги и позволяет производить сыры без кожуры, которые лучше подходят для высокой степени механизации (например, нарезки). При производстве сыров без кожуры буханки упаковывают в термоусадочную или безусадочную пленку с высокой проницаемостью CO2 (более 680 см3/м2/день при 23°C и 93% RH; Anonymous, 1991), но относительно низкой проницаемостью O2 (менее 100 см3/м2/день при 23°C и 93% RH) для предотвращения роста плесени на поверхности сыра. Если созревание в теплом помещении проводится для ускорения пропионовокислого брожения, сыры, обернутые пленкой, необходимо стабилизировать с помощью рам для сохранения их формы.
Сыры также могут быть покрыты эмульсиями виниловых полимеров, такими как Plasticoat. В этом случае относительная влажность воздуха должна быть выше, чем для сыров, созревающих в пластиковой пленке, что увеличивает риск роста плесени на поверхности или под покрытием. Поэтому эмульсии, используемые для покрытия, могут содержать противогрибковые ингредиенты, такие как натамицин или сорбиновая кислота.
Некоторые сыры швейцарского типа сначала созревают под полиэтиленовой пленкой, пока не завершится созревание в теплом помещении и не замедлится выделение CO2. Затем сыры разворачивают и покрывают воском или полимерной эмульсией.
Упаковка сыров пленками с разной проницаемостью CO2 может повлиять на образование глазков. На рис. 35.11 показаны рассчитанные средние объемы глазков сыров, упакованных четырьмя различными упаковочными пленками. У сыра, упакованного в пленку с самой высокой проницаемостью CO2 (P4), увеличение объема глазков происходит несколько медленнее.

Рис. 35.11 Развитие объема глазков во время созревания в теплом помещении с использованием пластиковых пленок P1, P2, P3, P4; пленка P1 имеет самую низкую проницаемость для CO2, а пленки P2, P3 и P4 становятся все более проницаемыми. Объем глазков был рассчитан по рентгеновским снимкам с использованием “pixel” в качестве меры длины для оценки радиуса отдельных глазков. (Перепечатано из Kraggerud, H., Wold, J.P., Hoy, M. and Abrahamsen, R.K., 2009. X-ray images for the control of eye formation in cheese. Int. J. Dairy Technol. 62, 147-153.).
СОЗРЕВАНИЕ
Формирование глазков

Текстура сыра и образование глазков - очень важные параметры качества для производителей, розничных продавцов и потребителей. В пределах одного сорта сыра допустимы лишь небольшие различия. Существует тесная взаимосвязь между текстурой, открытостью, вкусом и способностью к созреванию. Оценка образования глазков традиционно проводится путем прослушивания типа звука при постукивании по поверхности сыра специальным молоточком, визуального осмотра небольшого цилиндра сыра с помощью пробоотборника или просмотра разрезанной на половинки части сыра. Однако эти методы сильно зависят от опыта испытателя и не очень точны. Более новые методы неинвазивного получения изображений с использованием рентгеновских лучей (Akkerman et al., 1989; Blanc and Hättenschwiler, 1973; Kraggerud et al., 2009), ультразвука (Albrecht et al., 1998; Conde et al., 2008; Eskelinen et al., 2007), рентгеновской компьютерной томографии (CT) (Abrahamsen et al., 2006; Guggisberg et al.., 2013; Schuetz et al., 2013; Strand, 1985) и магнитно-резонансная томография (MRI) (Huc et al., 2013a; Musse et al., 2014; Rosenberg et al., 1992) применяются для исследования формирования глазков в сыре, особенно в отношении кинетики роста, размера, количества, пространственного распределения и дефектов глазков сыра. Размер, количество, форма и распределение глазков в сыре швейцарского типа являются важнейшими показателями качества. Образование глазков в сыре зависит в основном от времени, количества и интенсивности выделения и диффузии CO2, наличия ядер глазков, а также от текстуры и температуры сырного тела.
 CO2 в сырной массе сыров Аппенцеллер (N = 182), Эмменталер PDO (N = 159) и Грюйер PDO (N = 29)")
Рис. 35.12 Боксплот («Ящик с усами») CO2 в сырной массе сыров Аппенцеллер (N = 182), Эмменталер PDO (N = 159) и Грюйер PDO (N = 29); (неопубликованные результаты Agroscope, Берн, Швейцария). Медиана находится между нижним и верхним квартилями; усы показывают вариабельность за пределами нижнего и верхнего квартилей.
В сырах швейцарского типа образование глазков происходит в основном в результате выделения CO2 при пропионовокислом брожении во время созревания в теплом помещении (около 60 дней при 20-24°C для Эмменталера PDO). Однако меньшее количество CO2 образуется уже во время молочнокислого брожения и при распаде цитрата. Как только образуется достаточное количество глазков (примерно через 70 дней после производства), производство CO2 снижается путем хранения сыра при более низких температурах (10-13°C). По мнению Martley и Crow (1996), насыщение сырной матрицы CO2 (>18-36 ммоль/кг) является необходимым условием для образования глазков. Это может быть реализовано только при высокой скорости производства CO2 и относительно низкой скорости диффузии CO2 из сыра. Сравнение содержания CO2 в сырной массе сыров Эмменталер PDO (большие глазки), Аппенцеллер (маленькие круглые глазки, CO2 образуется в результате гетероферментативного молочнокислого брожения) и Грюйер PDO (глазки отсутствуют) показано на рис. 35.12.
Количество и распределение глазков также сильно зависит от технологической обработки, такой как фильтрация, центрифугирование, бактофугирование или микрофильтрация сырного молока. Центрифугирование и тепловая обработка молока или применение вакуума во время формования или прессования сыра проводятся для получения меньшего количества глазков. В сырах, произведенных из микрофильтрованного молока, количество глазков обычно значительно меньше (Fragnière and Schafroth, 2004). Столетие назад Clark (1917) предположил, что образование глазков в сыре происходит в "благоприятных местах" и происходит по принципу кристаллизации из пересыщенного раствора, когда начало роста кристаллов обязательно инициируется небольшими "семенами" или "неровностями" и когда размер конечных кристаллов зависит от количества добавленных семян. Азот из молока, CO2 из заквасок, твердые микрочастицы, небольшие механические отверстия и микроскопические пузырьки воздуха, попавшие в творог, были предложены в качестве ядер, в которых CO2 диссоциирует из растворимого состояния в теле сыра (Polychroniadou, 2001; Martley and Crow, 1996).
Недавние результаты показывают, что следовые количества сенного порошка, добавленные в микрофильтрованное молоко, вызывают образование глазков во время созревания сыра (рис. 35.13; Guggisberg et al., 2015). Микроскопическое исследование микрочастиц сенного порошка показало, что капиллярные структуры, присутствующие в растительных тканях из листьев или стеблей, вероятно, являются эффективными структурными элементами, которые функционируют как ядра глазков и обеспечивают образование глазков во время созревания сыра. Возможный захват воздуха в таких капиллярах обеспечивает диффузию СО2 из тела сыра в микрочастицы (рис. 35.14).
Многомасштабное исследование глазков в сыре швейцарского типа было проведено Huc et al. (2013a,b, 2014), чтобы проверить связь между кинетикой роста глазков и микроструктурой сыра вокруг глазков. Методы MRI сочетались с тремя микроскопическими методами [световая микроскопия, CLSM и SEM (от макро- до микромасштаба)] для определения организации жировой и белковой сети в матрице сыра вокруг глазков. Оказалось, что творожные зерна сильно растянуты вокруг отверстий (рис. 35.15).

Рис. 35.13 Выявленные с помощью рентгеновской компьютерной томографии сырные глазки в экспериментальных эмментальских сырах 45-дневной выдержки. Четыре образца сыра были произведены с разным количеством сухого порошкообразного сена (0,125-1,000 мг/90 л молока). Цвет глаз зависит от их размера. (Адаптировано из Guggisberg, D., Schuetz, P., Winkler, H., Amrein, R., Jakob, E., Fröhlich-Wyder, M.T., Irmler, S., Bisig, W., Jerjen, I., Plamondon, M., Hofmann, J., Flisch, A., Wechsler, D., 2015. Mechanism and control of the eye formation in cheese. Int. Dairy J. 47, 118-127).
, которые могут функционировать как ядра глазков в сыре")
Рис. 35.14 На микрофотографии порошкообразного сена видны структуры с капиллярными элементами (стрелки), которые могут функционировать как ядра глазков в сыре. Попадание воздуха в такие капилляры обеспечивает диффузию СО2 из сырного тела в микрочастицы. Последующий рост полостей в конечном итоге приводит к образованию видимых глазков в сыре. (Перепечатано из Guggisberg, D., Schuetz, P., Winkler, H., Amrein, R., Jakob, E., Fröhlich-Wyder, M.T., Irmler, S., Bisig, W., Jerjen, I., Plamondon, M., Hofmann, J., Flisch, A., Wechsler, D., 2015. Mechanism and control of the eye formation in cheese. Int. Dairy J. 47, 118-127).

Рисунок 35.15 Наблюдения глазков в сыре: (А) левая сторона отверстия при световой микроскопии (x4), (B) верхняя сторона отверстия при световой микроскопии (x4) и (C) схематическое расположение сырной массы вокруг глазков. (Перепечатано из Huc, D., Moulin, G., Mariette, F., Michon, C., 2013b. Investigation of curd grains in Swiss-type cheese using light and confocal laser scanning microscopy. Int. Dairy J. 33, 10-15.).
Текстура
Текстура обозначает структуру и консистенцию сырного тела. Мягкая и эластичная текстура имеет решающее значение для правильного формирования глазков. Большое количество кальция, связанного с казеином, что является результатом высокого значения рН при отделении сыворотки и после молочнокислого брожения (рН 5,20-5,35), очень важно для "длинной" эластичной текстуры (Lucey and Fox, 1993; Lucey et al., 2003). На рис. 35.16 показано, что содержание кальция в Эмменталере PDO относительно высокое по сравнению с другими сортами сыра. Famelart et al. (2002) также обнаружили, что содержание кальция во французском сыре Эмменталь составляет около 10 г/кг. Сыр Раклетт, который употребляется в основном в расплавленном виде, имеет довольно низкое содержание кальция (Fröhlich-Wyder et al., 2009; Sieber, 2012). Чеддер, плавящиеся свойства которого сопоставимы с сыром раклет, также имеет содержание кальция около 6-7 г/кг (Lucey and Fox, 1993; Lucey et al., 2005). Кроме того, содержание казеин-ассоциированного кальция в эмментальских сырах выше, чем в других сортах, что является следствием повышенного рН во время созревания (Lucey and Fox, 1993). Этот структурообразующий кальций необходим для эластичной текстуры. Сравнение реологических параметров твердых сортов сыра Эмменталер PDO и Грюйер PDO и полутвердого сорта Аппенцеллер показывает, что сыр Эмменталер PDO на самом деле является самым эластичным сортом, а деформация при разрушении самая высокая (рис. 35.17).
, Грюйере PDO (N = 204) и Раклетте (N = 1257)")
Рис. 35.16 Боксплот содержания кальция в Эмменталере PDO (N = 289), Грюйере PDO (N = 204) и Раклетте (N = 1257); (неопубликованные результаты Agroscope, Берн, Швейцария). Медиана находится между нижним и верхним квартилями; усы показывают изменчивость за пределами нижнего и верхнего квартилей.
, Emmentaler PDO (N = 83) и Gruyère PDO (N = 66)")
Рисунок 35.17 Боксплоты деформации при разрушении, полученные в результате испытания на одноосное сжатие для Appenzeller (N = 48), Emmentaler PDO (N = 83) и Gruyère PDO (N = 66); (неопубликованные результаты Agroscope, Берн, Швейцария). Медиана находится между нижним и верхним квартилями; усы указывают на вариабельность за пределами нижнего и верхнего квартилей.
Формирование аромата и вкуса

Основные первичные реакции при производстве сыра включают: (1) ферментацию лактозы с помощью МКБ, (2) ферментацию цитрата с помощью ФГЛ и (3) ферментацию лактата с помощью ПКБ. Образующиеся при этом метаболиты — лактат, пропионат, уксусная кислота, муравьиная кислота, ацетоин, диацетил — вносят значительный вклад в типичный вкус сыров швейцарского типа. Лактат вырабатывается в течение первых 24 часов производства, тогда как другие метаболиты, главным образом короткоцепочечные жирные кислоты, образуются в период пребывания в теплом помещении.
Протеолиз считается важным процессом созревания, способствующим формированию аромата за счет высвобождения свободных аминокислот, которые в дальнейшем катаболизируются до летучих ароматических соединений. Наиболее быстрая скорость накопления аминокислот наблюдается в период теплого помещения (Chen et al., 2009; Ji et al., 2004; Kocaoglu-Vurma et al., 2008). Основной вклад вносят молочнокислые бактерии, а на параметры протеолиза присутствие ПКБ существенно не влияет (Rehn et al., 2011; Thierry et al., 2004a; Wyder et al., 2001). Напротив, присутствие ПКБ влияет на аминокислотный профиль. Аспартат-положительные ПКБ преобразуют аспартат через фумарат в янтарную кислоту и NH+, которые являются мощными вкусовыми компонентами (рис. 35.4). Вместе с глутаминовой и пропионовой кислотами янтарная кислота способствует появлению вкуса умами в сыре швейцарского типа (Drake et al., 2007). Кроме того, ПКБ обладают высокой способностью превращать аминокислоты с разветвленной цепью - лейцин, изолейцин и валин - в карбоновые кислоты и спирты, причем изолейцин является наиболее предпочтительной катаболизируемой аминокислотой (Thierry et al., 2004a). Изолейцин превращается в основном в 2-метилмасляную кислоту, а лейцин - в изовалериановую кислоту (3-метилмасляную кислоту), причем концентрация в сыре в 3-10 раз выше, чем в отсутствие ПКБ (Thierry et al., 2004b) (рис. 35.18). Ниже приводится описание вкусовых характеристик изовалериановой кислоты: прогорклый, сладковатый, сырный, гнилостный (Smit et al., 2005).
Липолиз, по общему признанию, необходим для придания сыру типичного вкуса швейцарского типа. Гидролиз глицеридов приводит к высвобождению свободных жирных кислот (СЖК), которые могут быть предшественниками других вкусовых соединений в сыре. Источниками липолиза в сыре швейцарского типа являются бактериальные липазы и липопротеиновая липаза из молока, которая термолабильна, поэтому ее активность снижается при температуре приготовления свыше 50°C или устраняется при пастеризации сырного молока. Молочнокислые бактерии обычно проявляют лишь ограниченную липолитическую активность. В отличие от них, ПКБ обладают высокой штаммозависимой липолитической активностью: в 10-100 раз большей, чем молочнокислые бактерии. Количество свободных жирных кислот в сырах швейцарского типа варьируется от 2 до 7 г/кг, однако более высокое содержание приводит к ухудшению вкуса (Chamba and Perreard, 2002). Выделение свободных жирных кислот начинается в теплом помещении одновременно с ростом ПКБ (Chamba and Perreard, 2002) и продолжается в течение последующего периода холодного созревания (Dherbecourt et al., 2010; Lopez et al., 2006) (табл. 35.2). Предполагается, что липолиз под действием P. freudenreichii происходит за счет активности внеклеточных эстераз, а не внутриклеточных эстераз, высвобождаемых из лизированных клеток (Abeijon Mukdsi et al., 2014; Dherbecourt et al., 2010). Поскольку ПКБ являются основным агентом липолиза в сырах швейцарского типа, а липолитическая активность сильно зависит от штамма, важно учитывать этот критерий при разработке и выборе новых культур ПКБ.
Большинство летучих соединений образуется в результате гликолиза, протеолиза, липолиза и катаболизма образующихся метаболитов во время созревания и включает летучие короткоцепочечные кислоты (рис. 35.6), спирты, кетоны, альдегиды, эфиры, лактоны, алканы, ароматические углеводороды и различные серо- и азотсодержащие соединения (табл. 35.3). Warmke et al. (1996) оценили следующие вещества как мощные вкусовые соединения: уксусная, пропионовая, молочная, янтарная и глутаминовая кислоты, каждая в свободной форме и/или в виде аммонийных, натриевых, калиевых, магниевых и кальциевых солей, а также соответствующих хлоридов и фосфатов. Именно соли пропионовой кислоты в основном ответственны за сладковатый вкус сыров швейцарского типа, а образование эфиров и органических кислот под действием ПКБ считается вкладом в типичный аромат (Thierry et al., 2004a, 2004b, 2005).
Эфиры отвечают за фруктовые ароматы в сыре; эфиры пропионата являются наиболее распространенными эфирами в сыре швейцарского типа (табл. 35.3). Альдегиды и кетоны отвечают за многие известные вкусы и запахи, например, ванили, корицы, мяты и миндаля. В сыре швейцарского типа они обусловливают приятные фруктовые и солодовые нотки.
, использованных при производстве небольших экспериментальных сыров швейцарского типа, на количество метилмасляной кислоты, образующейся после 28 дней созревания в теплом помещении")
Рисунок 35.18 Влияние комбинации штаммов заквасок (LH: Lb. helveticus, LL: Lb. delbrueckii ssp. lactis и ITGP: P. freudenreichii), использованных при производстве небольших экспериментальных сыров швейцарского типа, на количество метилмасляной кислоты, образующейся после 28 дней созревания в теплом помещении. Простые столбики и столбики с индикаторами ошибок показывают средние значения и стандартные отклонения для трех экземпляров сыра. Значения с одинаковыми буквами существенно не различаются согласно тесту LSD (P < 0,05). (Перепечатано из Thierry, A., Richoux, R. and Kerjean, J.R., 2004b. Isovaleric acid is mainly produced by Propionibacterium freudenreichii in Swiss cheese. Int. Dairy J. 14, 801-807).
Таблица 35.2 Концентрация свободных жирных кислот (СЖК) в процессе созревания сыра Эмменталь a
|
Жирная кислота
|
Концентрация отдельных СЖК во время созревания (мг/г жира)
|
|||||
|
После варки
|
Конец хранения
при 12°C
|
Конец хранения
при 21°C
|
8 дней в помещении
при 4°C
|
Конец созревания
|
Statb
|
|
|
C4:0
|
321 ± 1
|
501 ± 1
|
3702 ± 14
|
4152 ± 58
|
4933 ± 10
|
***
|
|
C6:0
|
141 ± 3
|
212 ± 6
|
1433 ± 2
|
1423 ± 1
|
1423 ± 3
|
***
|
|
C8:0
|
221 ± 0
|
251 ± 2
|
1192 ± 5
|
1232 ± 2
|
1182 ± 7
|
***
|
|
C10:0
|
971 ± 3
|
1001 ± 8
|
3502 ± 5
|
4023 ± 27
|
3632 ± 15
|
***
|
|
C12:0
|
1061 ± 8
|
1121 ± 6
|
4612 ± 18
|
5573 ± 37
|
4742 ± 7
|
***
|
|
C14:0
|
2071 ± 7
|
2271 ± 11
|
13842 ± 74
|
17663 ± 109
|
13752 ± 81
|
***
|
|
C16:0
|
6991 ± 25
|
7051 ± 32
|
49652 ± 240
|
61873 ± 424
|
50012 ± 100
|
***
|
|
C18:0
|
2741 ± 47
|
2721 ± 36
|
12382 ± 41
|
15743 ± 128
|
12962 ± 41
|
***
|
|
C18:1
|
6641 ± 21
|
7081 ± 59
|
35322 ± 89
|
38883 ± 138
|
35972 ± 69
|
***
|
|
C18:2
|
661 ± 3
|
961 ± 29
|
2872 ± 6
|
3443 ± 25
|
3423 ± 20
|
***
|
|
C18:3
|
411 ± 3
|
551 ± 19
|
1222 ± 4
|
1483 ± 12
|
1553 ± 12
|
***
|
|
Σ (C4:0−C8:0)
|
681 ± 4
|
962 ± 5
|
6323 ± 22
|
6804 ± 60
|
7535 ± 18
|
***
|
|
Σ*(C10:0−C14:0)
|
4101 ± 18
|
4391 ± 25
|
21952 ± 96
|
27253 ± 172
|
22122 ± 71
|
***
|
|
Σ (C16:0−C18:3)
|
17441 ± 99
|
18361 ± 173
|
101442 ± 375
|
121413 ± 725
|
103912 ± 173
|
***
|
|
Σ (C4:0−C18:3)
|
22221 ± 126
|
23711 ± 193
|
129712 ± 491
|
155463 ± 957
|
133562 ± 167
|
***
|
(Перепечатано из Lopez, C., Maillard, M.B., Briard-Bion, V., Camier, B., Hannon, J.A., 2006. Lipolysis during ripening of Emmental cheese considering organization of fat and preferential localization of bacteria. J. Agric. Food Chem. 54, 5855-5867.)
Таблица 35.3 Влияние трех штаммов Propionibacterium freudenreichii (P17, P23 и P14) на концентрацию нейтральных летучих соединений в сырах
|
RTa (min)
|
Соединение
(нг/г)b
|
QIc
|
На конец хранения
в тепле d
|
После 2 месяцев хранения
при 4°C d
|
||||||||
|
Ctrle
|
P17
|
P23
|
P14
|
Statf
|
Ctrle
|
P17
|
P23
|
P14
|
Statf
|
|||
|
|
Спирты
|
|||||||||||
|
7.92
|
2-Бутанол
|
45
|
nd
|
nd
|
nd
|
nd
|
—
|
11.8b
|
4.0a
|
4.9a
|
3.3a
|
***
|
|
11.90
|
2-пентанол
|
45
|
3.9b
|
0.6a
|
0.8a
|
0.7a
|
*
|
23.3b
|
0.6a
|
5.0a
|
0.1a
|
*
|
|
13.63
|
3-Метилбутанол †
|
70
|
26.5a
|
31.5a
|
24.2a
|
64.0b
|
**
|
99.6
|
50.5
|
61.6
|
83.2
|
NS
|
|
13.75
|
2-Метилбутанол
|
57
|
3.4a
|
70.2b
|
77.6b
|
319.1c
|
***
|
11.7a
|
126.0c
|
77.9b
|
312.1d
|
***
|
|
|
Альдегиды
|
|||||||||||
|
6.29
|
2-Метилпропаналь
|
72
|
1.6
|
0.9
|
1.3
|
1.3
|
NS
|
4.7b
|
1.3a
|
1.8a
|
2.6a
|
**
|
|
10.00
|
3-Метилбутаналь †
|
TIC
|
8.5b
|
2.2a
|
3.1a
|
4.5a
|
***
|
8.2b
|
2.9a
|
3.2a
|
4.0a
|
**
|
|
10.42
|
2-Метилбутаналь
|
TIC
|
2.7a
|
17.3b
|
18.4b
|
51.7c
|
***
|
3.0a
|
14.6b
|
15.5b
|
29.2c
|
**
|
|
11.84
|
Пентанал #
|
44
|
2.1b
|
1.2a
|
1.0a
|
1.1a
|
**
|
2.5b
|
1.2a
|
1.5a
|
1.2a
|
*
|
|
|
Эфиры
|
|||||||||||
|
9.02
|
Метилпропионат #
|
88
|
0.0a
|
3.7b
|
3.0b
|
3.8b
|
***
|
0.0a
|
15.7b
|
14.0b
|
15.7b
|
***
|
|
12.51
|
Этилпропионат †
|
TIC
|
0.0a
|
27.6b
|
25.3b
|
27.5b
|
***
|
0.3a
|
195.5b
|
128.9b
|
158.9b
|
***
|
|
12.64
|
n-пропилацетат
|
43
|
0.2
|
0.4
|
0.9
|
0.3
|
NS
|
0.2a
|
1.6b
|
0.7a
|
0.4a
|
*
|
|
14.40
|
Изопропил-пропионат #
|
75
|
nd
|
nd
|
nd
|
nd
|
—
|
0.0a
|
0.4a
|
0.7b
|
0.4a
|
*
|
|
16.79
|
Пропилпропионат
|
57
|
0.02
|
0.05
|
0.01
|
0.06
|
NS
|
0.0a
|
1.0b
|
1.4b
|
1.8b
|
**
|
|
|
Кетоны
|
|||||||||||
|
7.29
|
2,3-Бутандион †
|
86
|
387
|
197
|
216
|
226
|
NS
|
494b
|
181a
|
284a
|
227a
|
**
|
|
7.60
|
2-Бутанон
|
72
|
195a
|
278a
|
293a
|
429b
|
*
|
305a
|
411a
|
376a
|
574b
|
*
|
|
11.74
|
2,3-Пентанедион #
|
100
|
1.9a
|
12.1b
|
8.0ab
|
10.3b
|
*
|
3.3a
|
13.5b
|
14.6b
|
10.6b
|
**
|
|
13.77
|
4-Метил-2-пентанон
|
43
|
1.5a
|
1.8a
|
2.3a
|
5.0b
|
***
|
2.1a
|
3.3a
|
3.5ab
|
5.0b
|
**
|
|
15.95
|
2-Гексанон
|
TIC
|
3.4a
|
9.9ab
|
6.1a
|
13.3b
|
*
|
8.6
|
12.6
|
8.8
|
9.8
|
NS
|
|
20.14
|
2-Гептанон †
|
TIC
|
142.7
|
104.6
|
105.6
|
105.8
|
NS
|
235.2b
|
111.1a
|
138.8a
|
124.3a
|
*
|
|
22.53
|
6-Метил-2-гептанон #
|
58
|
0.0
|
0.1
|
0.1
|
0.1
|
NS
|
0.0a
|
0.4b
|
0.5b
|
0.6b
|
***
|
|
23.01
|
5-Метил-2-гептанон #
|
43.58
|
0.0a
|
1.0bc
|
1.4c
|
0.7b
|
**
|
0.0a
|
0.8b
|
1.0b
|
0.7b
|
***
|
|
23.95
|
2-октанон #
|
58
|
1.7
|
3.2
|
2.2
|
5.2
|
NS
|
2.4a
|
4.2b
|
2.5ab
|
4.4b
|
*
|
|
|
S-содержащие соединения
|
|||||||||||
|
23.65
|
Диметил трисульфид
|
126
|
1.1b
|
0.3a
|
0.5a
|
0.4a
|
*
|
0.5
|
0.3
|
0.4
|
0.6
|
NS
|
(Перепечатано из Thierry, A., Maillard, M.B., Richoux, R., Kerjean, J.R. and Lortal, S. (2005). Propionibacterium freudenreichii strains quantitatively affect production of volatile compounds in Swiss cheese. Lait 85, 57-74).
ДЕФЕКТЫ СЫРА
Дефекты вкуса

Прогорклый привкус в сырах швейцарского типа чаще всего вызван чрезмерным липолизом сырого молока. Сыр, произведенный из пастеризованного молока, обычно подвергается меньшему липолизу, чем сыр из сырого молока, поскольку в первом случае липопротеиновая липаза деактивируется. Тем не менее термостойкие микробные липазы, присутствующие в молоке, хранившемся несколько дней, могут вызывать прогорклый привкус даже у сыра, изготовленного из пастеризованного молока.
Жгучий вкус
Перченый или жгучий вкус в сырах швейцарского типа чаще всего связан с присутствием гистамина в концентрациях, превышающих 200 мг/кг. В сыре Lactobacillus parabuchneri обычно, если не исключительно, отвечает за образование гистамина (Ascone et al., 2016; Fröhlich-Wyder et al., 2013). Жизнеспособное количество L. parabuchneri в молоке - вплоть до 100 КОЕ/мл - может провоцировать сильную выработку гистамина во время созревания. Сыры из сырого молока с температурой приготовления ниже ∼52°C особенно склонны к образованию гистамина, что всегда связано с дополнительным выделением CO2 в результате декарбоксилирования гистидина.
Горький вкус
В зависимости от местной молочной флоры, заквасочных культур и типа используемых коагулянтов, гидрофобные пептиды, полученные из казеинов, например, бета-казеин, могут накапливаться в сыре во время созревания и вызывать горький вкус. В сыре Эмменталер PDO слегка горький вкус иногда ощущается в возрасте 3 месяцев, но редко в сырах, созревающих в течение 8 месяцев и более. Это явление можно объяснить высокой пептидолитической активностью сыра, приводящей к деградации горьких пептидов.
Дефекты текстуры и консистенции

Чрезмерный протеолиз приводит к образованию слишком мучнистого и хрупкого тела, что делает сыр более склонным к образованию щелей и других дефектов глазков. Кроме того, когда большое количество казеина распадается до низкомолекулярных соединений, в сыре накапливаются плохо растворимые свободные аминокислоты, такие как тирозин. Это явление особенно ярко выражено в зоне около кожуры и чаще всего вызвано слишком быстрым охлаждением сырных пластов во время прессования, что замедляет молочнокислое брожение во внешней зоне сыров. Как следствие, может развиться бактериальная флора, которая протеолитически более активна, чем флора в центре буханки. Это обычно приводит к таким дефектам сыра, как мучнистое и твердое тело, резкий вкус или появление белого цвета под кожурой. Если протеолиз слишком низок, консистенция тела становится "длинной" или резиновой; иногда также появляются неровные отверстия.
Твердая консистенция
Консистенция сыра в основном зависит от содержания воды и жира, а также от температуры плавления жира. Консистенция сыров, произведенных в сезон кормления сеном (зимой), обычно более твердая, чем у сыров, произведенных в сезон кормления травой (летом), из-за более высокой температуры плавления жира. Опыт показывает, что сыры с более твердым телом более склонны к дефектам глаз. Поэтому некоторые производители сыра рекомендуют фермерам скармливать скоту семена масличных культур с высоким содержанием олеинового масла (т.е. масла с высоким содержанием олеиновой (омега-9) кислоты) в зимний период.
Дефектное формирование глазков
Дефекты, вызванные клостридиями
 Clostridium tyrobutyricum и (B) Clostridium sporogenes")
Рисунок 35.19 Дефекты сыра, вызванные (A) Clostridium tyrobutyricum и (B) Clostridium sporogenes (Agroscope, Берн, Швейцария)
Сыр швейцарского типа особенно подвержен дефектам сыра, вызванным клостридиями (Clostridium). Причинами являются относительно высокий уровень pH в сыре при засолке из-за промывки творога, созревание в теплом помещении в течение нескольких недель, большие сырные батоны и, как следствие, медленная диффузия соли в сырную массу. Чаще всего наблюдается маслянокислое брожение (также известное как «позднее вспучивание»), вызываемое C. tyrobutyricum (рис. 35.19 а). Кроме того, Clostridium sporogenes, обладающие сильным протеолитическим действием, могут вызывать появление белых пятен или ямок и гнилостный запах в сыре швейцарского типа (рис. 35.19 B). Аналогичный дефект может возникнуть на поверхности сыров швейцарского типа, созревших под полиэтиленовой пленкой (Bergère and Le noir, 2000). Clostridium beijerinckii также может влиять на формирование глазков и уровень масляной кислоты в сыре Эмменталь (Le Bourhis и др., 2007).
Дефект расщепления и вторичная ферментация

Рис. 35.20 Пример сыра пониженного качества швейцарского типа с содержанием гистамина 630 мг/кг, путресцина 480 мг/кг и тирамина 505 мг/кг, демонстрирующий сильное образование трещин в результате дополнительного выделения CO2 при декарбоксилировании соответствующих свободных аминокислот (Agroscope, Берн, Швейцария)
Из-за образования большого количества CO2 в течение нескольких недель (обычно > 500 мл CO2 на кг сыра) сыры швейцарского типа особенно склонны к образованию трещин, если эластичность сырного тела недостаточна. Это может произойти, если во время молочнокислого брожения pH опускается ниже 5,20, протеолиз протекает слишком интенсивно и/или консистенция сырного тела слишком твердая. Чрезмерный протеолиз также может привести к дополнительному выделению CO2, поскольку свободные аминокислоты могут быть декарбоксилированы микроорганизмами, присутствующими в сыре. Кроме того, стимулируется работа ПКБ, поскольку свободные аспарагин и аспартат утилизируются аспартат-положительными ПКБ (Fröhlich-Wyder et al., 2002). Чрезмерное выделение CO2 в сыре швейцарского типа на более поздней стадии созревания (например, после теплого помещения) с одновременным образованием больших глазков и трещин или расслоений называется поздней или вторичной ферментацией. Часто тело сыра не выдерживает давления газа, что приводит к появлению трещин или расслоений (рис. 35.20).
Образование редких (немногочисленных) глазков
")
Рис. 35.21 Малоглазковый сыр швейцарского типа с трещиной (Agroscope, Берн, Швейцария) (см. раздел "Цветные пластинки").
Размер, количество, форма и распределение глазков в сыре швейцарского типа являются важнейшими параметрами качества. Если количество глазков слишком мало или они неравномерно распределены в сырном пласте, нарезанные куски или ломтики могут иметь большие колебания в весе и быть частично лишенными глазков, тем самым не имея типичного внешнего вида сыра. Обработка молока, такая как фильтрация, бактофугирование или микрофильтрация, устраняет физические примеси (например, сенную пыль), присутствующие в сыром молоке. Это приводит к образованию редких глазков, поскольку такие примеси служат ядрами для образования глазков (Guggisberg et al., 2015; Polychroniadou, 2001). В малоглазковых сырах швейцарского типа средний размер глазков больше. Кроме того, сыры с редкими глазками более склонны к образованию трещин, чем сыры с оптимальными глазками (рис. 35.21). Недостаточное образование глазков также может быть связано с замедлением пропионовокислого брожения из-за низкого рН сыра (Turner et al., 1983) и/или недостаточной закваски культуры ПКБ. К другим факторам, замедляющим пропионовокислое брожение, относятся высокая температура ошпаривания (2-го нагревания), высокое содержание соли и высокий уровень меди в сыре.
Чрезмерное или неравномерное формирование глазков
Наличие мелких пузырьков воздуха в молоке (например, из-за эффекта Вентури в молокопроводах) может привести к нежелательному увеличению количества глазков в сыре. Кроме того, образование газа микроорганизмами, такими как колиформные бактерии или облигатно гетероферментативные молочнокислые бактерии, может привести к чрезмерному образованию глазков, особенно если активность закваски нарушена (O'Sullivan et al., 2016). Неравномерное распределение глазков в сырах швейцарского типа часто происходит при попадании воздуха в творог во время формования (рис. 35.22). Известно, что недостаточное прессование также провоцирует подобные дефекты.

Рис. 35.22 Пример чрезмерного образования глазков в результате захвата воздуха во время формования сырной массы в нижней части проиллюстрированного сыра (Agroscope, Берн, Швейцария).

Рис. 35.23 Пример неправильного формирования глазка в качественном сыре швейцарского типа пониженной категории (Agroscope, Берн, Швейцария) (см. раздел "Цветные пластинки").
Неправильное формирование глазков
Снижение рейтинга сыров с неправильными глазками (рис. 35.23) приводит к значительным финансовым потерям для производителя. Считается, что такие дефекты глазков связаны с недостаточным качеством текстуры сырного тела. Когда мы сталкиваемся с сырами с такой дефектной открытостью, консистенция сырного тела часто оказывается слишком твердой и сухой, а эластичность - слишком низкой. Помимо нормальных глазков, в одном и том же теле могут встречаться различные формы дефектов глазков, такие как ореховидные глазки, неправильно вложенные глазки и глазки, похожие на трещины, которые, по-видимому, имеют одно и то же происхождение. Считается, что в таких случаях сплавление сырных зерен является недостаточным. Однако трудно определить причины, ответственные за неправильное формирование глазков, поскольку микробиологические и технологические факторы способствуют недостаточной текстуре сыра, которая не позволяет образовываться нормальным глазкам.
ГИГИЕНИЧЕСКАЯ БЕЗОПАСНОСТЬ СЫРОВ ШВЕЙЦАРСКОГО ТИПА

Сыры Emmentaler PDO, Emmental de Savoie PGI и Emmental français est-central PGI должны производиться из сырого молока. Несмотря на интенсивные гигиенические мероприятия, полностью исключить заражение сырого молока патогенными микроорганизмами невозможно. Инфекционные заболевания молочных коров или загрязнение молока во время доения, хранения, транспортировки или переработки представляют потенциальную опасность. Это привело к дискуссиям о гигиенической безопасности сыров из сырого молока. Bachmann и Spahr (1995) и Spahr и Schafroth (2001) исследовали способность потенциально патогенных бактерий расти и выживать во время производства и созревания экспериментального эмментальского сыра, изготовленного из сырого молока (рис. 35.24). Они пришли к выводу, что гигиеническая безопасность традиционного эмментальского сыра сопоставима с сыром, произведенным из пастеризованного молока. На самом деле твердые сыры швейцарского типа имеют отличные показатели пищевой безопасности даже при изготовлении из сырого молока.

Рисунок 35.24 Поведение Aeromonas hydrophila (◊), Campylobacter jejuni (♦), Escherichia coli (∆), Listeria monocytogenes (▲), Pseudomonas aeruginosa (○), Salmonella typhimurium (○), Staphylococcus aureus (□) и Yersinia enterocolitica (■) во время производства и созревания швейцарского Эмменталера из сырого молока (показаны только данные партий с наибольшим выживанием). - - -, предел обнаружения. (Перепечатано из Bachmann, H.P., Spahr, U., 1995. The fate of potentially pathogenic bacteria in Swiss hard and semihard cheeses made from raw milk. J. Dairy Sci. 78, 476-483).
Снижение количества патогенов можно объяснить совместным антагонистическим действием заквасочной культуры, длительным выдерживанием сырного зерна при температуре выше 50°C, антимикробным действием молочной, уксусной и пропионовой кислот, а также созреванием в течение не менее 3 месяцев. Поскольку рост стафилококков подавляется вскоре после начала варки творога (сырного зерна), образование стафилококковых энтеротоксинов маловероятно. В сортах сыра швейцарского типа, которые производятся при температуре варки ниже 48°C, патогенные микроорганизмы, такие как Salmonella spp., L. monocytogenes, S. aureus и энтеропатогенная кишечная палочка, могут представлять риск для безопасности сыра. Поэтому при производстве таких сыров молоко обычно подвергается тепловой обработке.
Доп. информация о сыроделии:
- Пропионибактерии в сыроделии
- Сыроделие (подборка информации) + ссылки →
- Список PDF-информации о сыроделии
- Abeijon Mukdsi, M.C., Falentin, H., Maillard, M.B., Chuat, V., Medina, R.B., Parayre, S., Thierry, A., 2014. The secreted esterase of Propionibacterium freudenreichii has a major role in cheese lipolysis. Appl. Environ. Microbiol. 80, 751–756.
- Abrahamsen, R.K., Byre, O., Steinsholt, K., Strand, A.H., 2006. Jarlsberg Cheese History and Development. Tun Forlag, As, Norway.
- Akkerman, J.C., Walstra, P., Van Dijk, H.J.M., 1989. Holes in Dutch-type cheese. 1. Conditions allowing eye formation. Neth. Milk Dairy J. 43, 453–476.
- Albrecht, B., Ulmann, H.R., Bosset, J.O., 1998. Nachweis von Ris- sen in Käselaiben mit Ultraschall-Echografie. Dtsch. Milchwirt. 49, 1162–1163.
- Anonymous, 1991. Flexible wrappings (including laminates) for rindless cheese, Australian Standard AS2183; The Crescent, Hombush, NSW 2140, Standards Association of Australia.
- Anonymous, 2012. Emmentaler (GUB). Federal Office for Agriculture FOAG, Bern, Switzerland.
- Ardö, Y., Varming, C., 2010. Bacterial influence on characteristic flavour of cheeses made with mesophilic dlstarter. Aust. J. Dairy Technol. 65, 153–158.
- Ascone, P., Maurer, J., Haldemann, J., Irmler, S., Berthoud, H., Portmann, R., Fröhlich-Wyder, M.T., Wechsler, D., 2016. Prevalence and diversity of histamine-forming Lactobacillus parabuchneri strains in raw milk and cheese – A case study. Int. Dairy J. doi: 10.1016/j.id- airyj.2016.11.012.
- Bachmann, H.P., Spahr, U., 1995. The fate of potentially pathogenic bacteria in Swiss hard and semihard cheeses made from raw milk. J. Dairy Sci. 78, 476–483.
- Baer, A., Ryba, I., 1999. Interactions between propionic acid bacteria and thermophilic lactic acid bacteria. Lait 79, 79–92.
- Beresford, T.P., Fitzsimons, N.A., Brennan, N.L., Cogan, T.M., 2001. Recent advances in cheese microbiology. Int. Dairy J. 11, 259–274.
- Bergère, J.L., Lenoir, J., 2000. Cheese manufacturing accidents and cheese defects. In: Eck, A., Gillis, J.-C. (Eds.), Cheesemaking—From Science to Quality Assurance. second ed. Lavoisier Publishing Inc, Paris cedex France.
- Bisig, W., Fröhlich-Wyder, M.T., Jakob, E., Wechsler, D., 2010. Comparison between Emmentaler PDO and generic Emmental cheese production in Europe. Aust. J. Dairy Technol. 65, 206–213.
- Blanc, B., Hättenschwiler, J., 1973. Neue Möglichkeiten für die Anwendung der Radiologie in der Käsereitechnologie. Schweiz. Milchwirt. Forschung 2, 1–15.
- Blasco, L., Kahala, M., Tupasela, T., Joutsjoki, V., 2011. Determination of aspartase activity in dairy Propionibacterium strains. FEMS 321, 10–13.
- Bouton, Y., Buchin, S., Duboz, G., Pochet, S., Beuvier, E., 2009. Effect of mesophilic lactobacilli and enterococci adjunct cultures on the final characteristics of a microfiltered milk Swiss-type cheese. Food Microbiol. 26, 183–191.
- Brendehaug, J., Langsrud, T., 1985. Amino acid metabolism in propionibacteria: resting cells experiments with four strains. J. Dairy Sci. 68, 281–289.
- Chamba, J.F., 2000. Emmental cheese: a complex microbial ecosystem. Consequences on selection and use of starters. Sci. Aliment. 20, 37–54.
- Chamba, J.F., Perreard, E., 2002. Contribution of propionic acid bacteria to lipolysis of Emmental cheese. Lait 82, 33–44.
- Chen, G., Kocaoglu-Vurma, N.A., Harper, W.J., Rodriguez-Saona, L.E., 2009. Application of infrared microspectroscopy and multivariate analysis for monitoring the effect of adjunct cultures during Swiss cheese ripening. J. Dairy Sci. 92, 3575–3584.
- Clark, W.M., 1917. On the formation of “Eyes” in Emmental cheese. J. Dairy Sci. 1, 91–113.
- Codex, 2014. Codex Standard for Emmental, Standard 269-1967, Established 1967, Revision 1 2007, Amendment 3 2013. Codex Alimenta- rius Commission, Joint FAO/WHO food standards programme.
- Conde, T., Mulet, A., Clemente, G., Benedito, J., 2008. Detection of internal cracks in manchego cheese using the acoustic impulse-response technique and ultrasounds. J. Dairy Sci. 91, 918–927.
- Crow, V.L., 1986. Metabolism of aspartate by Propionibacterium freudenreichii subsp. shermanii: effect on lactate fermentation. Appl. Environ. Microbiol. 52, 359–365.
- Crow, V.L., Martley, F.G., Delacroix, A., 1988. Isolation and properties of aspartase-deficient variants of Propionibacterium freudenreichii subsp. shermanii and their use in the manufacture of Swiss-type cheese. New Zeal. J. Dairy Sci. 23, 75–85.
- Crow, V.L., Turner, K.W., 1986. The effect of succinate production on other fermentation products in Swiss-type cheese. New Zeal. J. Dairy Sci. 21, 217–227.
- Daly, D.F.M., McSweeney, P.L.H., Sheehan, J.J., 2010. Investigation of underlying factors in split and secondary fermentation defects in rindless block Swiss-type cheeses. Aust. J. Dairy Technol. 65, 214–216.
- Deutsch, S.M., Ferain, T., Delcour, J., Lortal, S., 2002. Lysis of lysogenic strains of Lactobacillus helveticus in Swiss cheeses and first evidence of concomitant Streptococcus thermophilus lysis. Int. Dairy J. 12, 591–600.
- Dherbecourt, J., Bourlieu, C., Maillard, M.B., Aubert-Frogerais, L., Richoux, R., Thierry, A., 2010. Time course and specificity of lipolysis in Swiss cheese. J. Agr. Food Chem. 58, 11732–11739.
- Drake, S.L., Whetstine, M.E.C., Drake, M.A., Courtney, P., Fligner, K., Jenkins, J., Pruitt, C., 2007. Sources of umami taste in Cheddar and Swiss cheeses. J. Food Sci. 72, 360–366.
- Eskelinen, J.J., Alavuotunki, A.P., Haeggstrom, E., Alatossava, T., 2007. Preliminary study of ultrasonic structural quality control of Swiss-type cheese. J. Dairy Sci. 90, 4071–4077.
- European Commission, 2014. Agriculture and Rural Development, Quality policy, PDO-PGI-TSG—Geographical indications and traditional specialities.
- Falentin, H., Deutsch, S.M., Jan, G., Loux, V., Thierry, A., Parayre, S., Maillard, M.B., Dherbecourt, J., Cousin, F.J., Jardin, J., Siguier, P., Couloux, A., Barbe, V., Vacherie, B., Wincker, P., Gibrat, J.F., Gaillardin, C., Lortal, S., 2010. The complete genome of Propionibacterium freudenreichii CIRM-BIA1, a hardy actinobacterium with food and probiotic applications. PLoS One 5, e11748.
- Famelart, M.H., Le Graet, Y., Michel, F., Richoux, R., Riaublanc, A., 2002. Evaluation of the methods of measurement for functional properties of Emmental cheeses from the west of France. Lait 82, 225–245.
- FDA, 2014. Code of Federal Regulations Title 21 Food and Drugs, Chapter 1 Food and Drug Administration Department of health and human Services, Subchapter B: Food for human consumption, Part 133 Cheeses and related cheese products, Subpart B: Requirements for Specific Standardized Cheese and Related Products, Sec. 133.195 Swiss and emmentaler cheese. US Food and Drug Administration (cite 21CFR133) 2, revised 1st April 2013.
- Fessler, D., 1997. Characterisation of propionibacteria in Swiss raw milk by biochemical and molecular-biological methods. 12328 (PhD thesis). ETH Zürich, Switzerland.
- Fragnière, C., Schafroth, K., 2004. Neue Erkenntnisse zur Lochbildung in Käse aus mikrofiltrierter Milch. Schweiz. Milchztg 130, 7.
- Fröhlich-Wyder, M.T., Bachmann, H.P., Casey, M., 2002. Interaction between propionibacteria and starter/non-starter lactic acid bacteria in Swiss-type cheeses. Lait 82, 1–15.
- Fröhlich-Wyder, M.T., Eugster-Meier, E., 2005. Kann Grosslochkäse ohne Gärraumaufenthalt hergestellt werden? (Can Swiss type cheese be produced without a period in the fermentation room?). Agrarforschung 12, 410–415.
- Fröhlich-Wyder, M.T., Guggisberg, D., Badertscher, R., Wechsler, D., Wittwer, A., Irmler, S., 2013. The effect of Lactobacillus buchneri and Lactobacillus parabuchneri on the eye formation of semi-hard cheese. Int. Dairy J. 33, 120–128.
- Fröhlich-Wyder, M.T., Guggisberg, D., Wechsler, D., 2009. Influence of low calcium and low pH on melting characteristics of model Raclette cheese. Dairy Sci. Technol. 89, 463–483.
- Guggisberg, D., Fröhlich-Wyder, M.T., Irmler, S., Greco, M., Wechsler, D., Schuetz, P., 2013. Eye formation in semi-hard cheese: X-ray computed tomography as a non-invasive tool for assessing the influence of adjunct lactic acid bacteria. Dairy Sci. Technol. 93, 135–149.
- Guggisberg, D., Schuetz, P., Winkler, H., Amrein, R., Jakob, E., Fröhlich-Wyder, M.T., Irmler, S., Bisig, W., Jerjen, I., Plamondon, M., Hofmann, J., Flisch, A., Wechsler, D., 2015. Mechanism and control of the eye formation in cheese. Int. Dairy J. 47, 118–127.
- Huc, D., Mariette, F., Challois, S., Barreau, J., Moulin, G., Michon, C., 2013a. Multi-scale investigation of eyes in semi-hard cheese. Innov. Food Sci. Emerg. Technol. 24, 106–112.
- Huc, D., Moulin, G., Mariette, F., Michon, C., 2013b. Investigation of curd grains in Swiss-type cheese using light and confocal laser scanning microscopy. Int. Dairy J. 33, 10–15.
- Huc, D., Roland, N., Grenier, D., Challois, S., Michon, C., Mariette, F., 2014. Influence of salt content on eye growth in semi-hard cheeses studied using magnetic resonance imaging and CO2 production measurements. Int. Dairy J. 35, 157–165.
- Ji, T., Alvarez, V.B., Harper, W.J., 2004. Influence of starter culture ratios and warm room treatment on free fatty acid and amino acid in Swiss cheese. J. Dairy Sci. 87, 1986–1992.
- Jimeno, J., Lazaro, M.J., Sollberger, H., 1995. Antagonistic interactions between propionic acid bacteria and non-starter lactic acid bacteria. Lait 75, 401–413.
- Kerjean, J.R., Condon, S., Lodi, R., Kalantzopoulos, G., Chamba, J.F., Suomalainen, T., Cogan, T., Moreau, D., 2000. Improving the quality of European hard-cheeses by controlling of interactions between lactic acid bacteria and propionibacteria. Food Res. Int. 33, 281–287.
- Kocaoglu-Vurma, N.A., Harper, W.J., Drake, M.A., Courtney, P.D., 2008. Microbiological, chemical, and sensory characteristics of Swiss cheese manufactured with adjunct Lactobacillus strains using a low cooking temperature. J. Dairy Sci. 91, 2947–2959.
- Kraggerud, H., Wold, J.P., Hoy, M., Abrahamsen, R.K., 2009. X-ray images for the control of eye formation in cheese. Int. J. Dairy Technol. 62, 147–153.
- Langsrud, T., Reinbold, G.W., 1973. Flavor development and microbiology of Swiss cheese - a review. II. Starters, manufacturing processes and procedures. (Part 2 of 4). J. Milk Food Technol. 36, 531–542.
- Le Bourhis, A.G., Dore, J., Carlier, J.P., Chamba, J., Popoff, M.R., Tholo- zan, J.L., 2007. Contribution of C. beijerinckii and C. sporogenes in association with C. tyrobutyricum to the butyric fermentation in Emmental type cheese. Int. J. Food Microbiol. 113, 154–163.
- Lopez, C., Maillard, M.B., Briard-Bion, V., Camier, B., Hannon, J.A., 2006. Lipolysis during ripening of Emmental cheese considering organization of fat and preferential localization of bacteria. J. Agric. Food Chem. 54, 5855–5867.
- Lucey, J.A., Fox, P.F., 1993. Importance of calcium and phosphate in cheese manufacture: a review. J. Dairy Sci. 76, 1714–1724.
- Lucey, J.A., Johnson, M.E., Horne, D.S., 2003. Invited review: Perspec- tives on the basis of the rheology and texture properties of cheese. J. Dairy Sci. 86, 2725–2743.
- Lucey, J.A., Mishra, R., Hassan, A., Johnson, M.E., 2005. Rheological and calcium equilibrium changes during the ripening of Cheddar cheese. Int. Dairy J. 15, 645–653.
- Martley, F.G., Crow, V.L., 1996. Open texture in cheese: the contributions of gas production by microorganisms and cheese manufacturing practices. J. Dairy Res. 63, 489–507.
- McSweeney, P.L.H., Ottogalli, G., Fox, P.F., 2004. Diversity of cheese varieties: an overview. Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P. (Eds.), Cheese: Chemistry, Physics and Microbiology, Vol. 2, third ed. Major Cheese groups, Elsevier Ltd, London, pp. 1–22.
- Miks-Krajnik, M., Babuchowski, A., Bialobrzewski, I., 2013. Impact of physiological state of starter culture on ripening and flavour development of Swiss-Dutch-type cheese. Int. J. Dairy Technol. 66, 562–569.
- Musse, M., Challois, S., Huc, D., Quellec, S., Mariette, F., 2014. MRI method for investigation of eye growth in semi-hard cheese. J. Food Eng. 121, 152–158.
- Ojala, T., Laine, P.K.S., Ahlroos, T., Tanskanen, J., Pitkanen, S., Salusjarvi, T., Kankainen, M., Tynkkynen, S., Paulin, L., Auvinen, P., 2017. Functional genomics provides insights into the role of Propionibacterium freudenreichii ssp shermanii JS in cheese ripening. Int. J. Food Microbiol. 241, 39–48.
- O’Sullivan, D.J., McSweeney, P.L.H., Cotter, P.D., Giblin, L., Sheehan, J.J., 2016. Compromised Lactobacillus helveticus starter activity in the presence of facultative heterofermentative Lactobacillus casei DPC6987 results in atypical eye formation in Swiss-type cheese. J. Dairy Sci. 99, 2625–2640.
- Parizzi, L.P., Grassi, M.C., Llerena, L.A., Carazzolle, M.F., Queiroz, V.L., Lunardi, I., Zeidler, A.F., Teixeira, P.J., Mieczkowski, P., Rincones, J., Pereira, G.A., 2012. The genome sequence of Propionibacterium acidipropionici provides insights into its biotechnological and industrial potential. BMC Genom. 13, 562–582.
- Park, H.S., Reinbold, G.W., Hammond, W.S., Clark, Jr., W.S., 1967. Growth of propionibacteria at low temperatures. J. Dairy Sci. 50, 589–591.
- Perez Chaia, A., Pesce de Ruiz Holgado, A., Oliver, G., 1987. Interaction between Lactobacillus helveticus and Propionibacterium freudenreichii subsp. shermanii. Microbiol. Aliments Nutr. 5, 325–331.
- Piveteau, P., Condon, S., Cogan, T.M., 2000. Inability of dairy propionibacteria to grow in milk from low inocula. J. Dairy Res. 67, 65–71.
- Piveteau, P.G., Condon, S., Cogan, T.M., 1995. Interactions between lactic and propionic acid bacteria. Lait 75, 331–343.
- Piveteau, P.G., O’Callaghan, J., Lyons, B., Condon, S., Cogan, T.M., 2002. Characterisation of the stimulants produced by Lactobacillus helveticus in milk for Propionibacterium freudenreichii. Lait 82, 69–80.
- Polychroniadou, A., 2001. Eyes in cheese: a concise review. Milk Sci. Int.56, 74–77.
- Rehn, U., Vogensen, F.K., Persson, S.-E., Saeden, K.H., Nilsson, B.F., Ardö, Y., 2011. Influence of microflora on texture and contents of aminoacids, organic acids, and volatiles in semi-hard cheese made with dlstarter and propionibacteria. J. Dairy Sci. 94, 1098–1111.
- Reinbold, G.W., 1972. Swiss cheese varieties. Pfizer Inc, New York. Richoux, R., Faivre, E., Kerjean, J.R., 1998. Effect of NaCl content on lactate fermentation by Propionibacterium freudenreichii in small scale Swiss-type cheeses. Lait 78, 319–331.
- Richoux, R., Kerjean, J.R., 1995. Technological properties of pure propionibacteria strains: test in small scale Swiss-type cheese. Lait 75, 45–59.
- Rodriguez, L.M., Alatossava, T., 2008. Effects of copper supplement on growth and viability of strains used as starters and adjunct cultures for Emmental cheese manufacture. J. Appl. Microbiol. 105, 1098–1106.
- Rodriguez, L.M., Alatossava, T., 2010. Effects of copper on germination, growth and sporulation of Clostridium tyrobutyricum. Food Microbiol. 27, 434–437.
- Rodriguez, L.M., Ritvanen, T., Joutsjoki, V., Rekonen, J., Alatossava, T., 2011. The role of copper in the manufacture of Finnish Emmental cheese. J. Dairy Sci. 94, 4831–4842.
- Rosenberg, M., McCarthy, M., Kauten, R., 1992. Evaluation of eye formation and structural quality of Swiss-type cheese by magnetic-resonance-imaging. J. Dairy Sci. 75, 2083–2091.
- Samarzija, D., Antunac, N., Havranek, J.L., 2001. Taxonomy, physiology and growth of Lactococcus lactis: a review. Mljekarstvo 51, 35–48.
- Schuetz, P., Guggisberg, D., Jerjen, I., Fröhlich-Wyder, M.T., Hofmann, J., Wechsler, D., Flisch, A., Bisig, W., Sennhauser, U., Bachmann, H.P., 2013. Quantitative comparison of the eye formation in cheese using radiography and computed tomography data. Int. Dairy J. 31, 150–155.
- Scott, R., 1998. Cheesemaking Practice, third ed. Aspen Publishers, Inc, Gaithersburg, Maryland.
- Sheehan, J.J., Wilkinson, M.G., McSweeney, P.L.H., 2008. Influence of processing and ripening parameters on starter, non-starter and propionic acid bacteria and on the ripening characteristics of semi-hard cheeses. Int. Dairy J. 18, 905–917.
- Sieber, R., 2012. Zusammensetzung von Milch und Milchprodukten schweizerischer Herkunft. FAM-Information 538, 1–40.
- Sieber, R., Rehberger, B., Schaller, F., Gallmann, P., 2006. Technological aspects of copper in milk products and health implications of copper. ALP Sci. 493, 1–15.
- Smid, E.J., Lacroix, C., 2013. Microbe-microbe interactions in mixed culture food fermentations. Curr. Opin. Biotechnol. 24, 148–154.
- Smit, G., Smit, B.A., Engels, W.J.M., 2005. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 29, 591–610.
- Sollberger, H., Wyder, M.T., 2000. Propionsäurebakterien und fakultativ heterofermentative Laktobazillen. Schweiz. Milchztg 126, 5.
- Spahr, U., Schafroth, K., 2001. Fate of Mycobacterium aviumsubsp paratuberculosis in Swiss hard and semihard cheese manufactured from raw milk. Appl. Environ. Microbiol. 67, 4199–4205.
- Strand, A.H., 1985. Scanning of Jarlsberg using data-tomograph. Meieri- teknikk 39, 2–7.
- Thierry, A., Deutsch, S.M., Falentin, H., Dalmasso, M., Cousin, F.J., Jan, G., 2011. New insights into physiology and metabolism of Propionibacterium freudenreichii. Int. J. Food Microbiol. 149, 19–27.
- Thierry, A., Maillard, M.B., Herve, C., Richoux, R., Lortal, S., 2004a. Varied volatile compounds are produced by Propionibacterium freudenreichii in Emmental cheese. Food Chem. 87, 439–446.
- Thierry, A., Maillard, M.B., Richoux, R., Kerjean, J.R., Lortal, S., 2005. Propionibacterium freudenreichii strains quantitatively affect production of volatile compounds in Swiss cheese. Lait 85, 57–74.
- Thierry, A., Richoux, R., Kerjean, J.R., 2004b. Isovaleric acid is mainly produced by Propionibacterium freudenreichii in Swiss cheese. Int. Dairy J. 14, 801–807.
- Thierry, A., Salvat-Brunaud, D., Madec, M.N., Michel, F., Maubois, J.L., 1998. Affinage de l’emmental: dynamique des populations bactéri- ennes et évolution de la composition de la phase aqueuse. Lait 78, 521–542.
- Turgay, M., Irmler, S., Isolini, D., Amrein, R., Fröhlich-Wyder, M.T., Berthoud, H., Wagner, E., Wechsler, D., 2011. Biodiversity, dynamics, and characteristics of Propionibacterium freudenreichii in Swiss Emmentaler PDO cheese. Dairy Sci.Technol. 91, 471–489.
- Turner, K.W., Morris, H.A., Martley, F.G., 1983. Swiss-type cheese II. The role of thermophilic lactobacilli in sugar fermentation. New Zeal. J. Dairy Sci. Technol. 18, 117–123.
- Warmke, R., Belitz, H.D., Grosch, W., 1996. Evaluation of taste compounds of Swiss cheese (Emmentaler). Z. Lebensm. Unters. For. 203, 230–235.
- Weinrichter, B., Sollberger, H., Ginzinger, W., Jaros, D., Rohm, H., 2004. Adjunct starter properties affect characteristic features of Swiss-type cheeses. Food 48, 73–79.
- Wyder, M.T., Bosset, J.O., Casey, M.G., Isolini, D., Sollberger, H., 2001. Influence of two different propionibacterial cultures on the characteristics of Swiss-type cheese with regard to aspartate metabolism. Milk Sci. Int. 56, 78–81.